
陆地生态系统碳水通量特征研究进展
2022-02-06 06:35周军志李建柱陈伏龙
地球环境学报 2022年6期
张 婷,周军志,李建柱,陈伏龙,冯 平
1.天津大学 水利工程仿真与安全国家重点实验室,天津 300072
2.石河子大学 水利建筑工程学院,石河子 832003
陆地生态系统是地球陆地表面由陆生生物与其环境相互作用构成的统一整体,其按照植被特点和生态系统内部环境的不同,可分为森林、草地、农田、荒漠、湿地等子系统。陆地生态系统的碳循环、水循环是陆地生态系统最为重要的物质循环,它们相互依赖、相互制约,通过土壤 — 植物 — 大气系统的一系列能量转化、物质循环和水分传输过程紧密地耦联在一起,连接着地球系统各个圈层(地圈、大气圈和生物圈)(于贵瑞等,2004a),并对气候系统具有强烈的反馈作用。陆地生态系统碳水循环的过程在全球碳水循环中最复杂、受人类活动影响最大(李润东等,2020),其变化特征和响应机制在全球变暖背景下成为生态学研究的一个热点问题。
陆地生态系统碳水循环过程是地球陆地表层系统物质循环与能量交换的基本生物物理过程,只有深入研究陆地生态系统的生理过程机制和环境控制原理,才能探明生态系统的碳源汇机制和蒸散发过程(段玲玲等,2019)。与此同时,在我国提出2030年前CO2排放达到峰值,2060年前实现碳中和的背景下,增强陆地生态系统碳汇功能是实现碳达峰、碳中和目标的重要手段(杨元合等,2022)。不同生态系统碳水循环机理过程虽具有相似性,但对气候变化非常敏感,表现出显著的空间和时间变异性(He et al,2019)。本文通过查阅大量国内外文献资料,综述了CO2/ H2O通量研究的常用方法及其优势与不足,总结了我国不同陆地生态系统碳通量和水汽通量特征及其影响因子的研究进展,并对目前研究中存在的问题进行了简要分析,对进一步展开特定生态系统环境条件下的碳水循环过程研究具有重要意义。
1 碳水通量研究方法
通量观测和模型模拟是定量估计CO2/ H2O通量时空分布格局,理解碳水交换关键过程及其控制机制的两种常用方法(曹明奎等,2004)。
1.1 通量观测
陆地和大气系统间CO2通量与总初级生产力(GPP)、净初级生产力(NPP)、净生态系统生产力(NEP)概念是相对应的。通常条件下,在植被上部进行通量观测得到的CO2通量相当于生态系统的NEP。净生态系统碳交换量(NEE)主要受植物光合作用、生物呼吸作用和冠层空气中碳存储的共同影响,因此NEE可以表述为群落内碳储存通量和大气 — 生态系统界面的净CO2通量之和(于贵瑞和孙晓敏,2017)。在地势平坦、植被类型空间分布均匀的植被下垫面,涡度相关系统所观测的湍流涡度通量可以近似地认为等于净生态系统生产力。传统的碳通量观测主要是利用生态学调查和碳循环过程的测定方法。近代微气象观测技术的进步为植被 — 大气及土壤 — 大气界面的CO2/ H2O通量的直接测定提供了可能。涡度相关法作为评价植被群落水平CO2/ H2O通量最为有效的直接测定方法,已被微气象学家和生态学家广泛认同,被誉为目前检验各种测定方法和评价各类模型估算精度的标准方法(耿绍波等,2010)。
生态学调查和碳循环过程的测定方法包括基于生物量变化的估算法、基于碳平衡方程的估算法以及同化箱测定法(箱式法)(于贵瑞和孙晓敏,2017)。基于生物量变化的估算法,是利用NPP、NEP等可以直接反映生态系统的陆地 — 大气间NEE的概念,在一定假设条件下,利用生态系统生物量变化动态监测数据进行估算(Upchurch and Ritchie,1984)。基于碳平衡方程的估算法是通过利用各种测定技术对植物叶片的光合作用和呼吸作用、土壤微生物的呼吸通量、凋落物分解过程进行精确测定,来确定NEP以反映生态系统NEE。这两种测定方法只能适合于单叶或单株等十分有限的样品或样地,其测定数据的时间和空间代表性存在很多问题(于贵瑞和孙晓敏,2017)。土壤 — 大气界面测定生态系统呼吸通量主要采用扩散法和箱式法进行观测研究。扩散法利用土壤空气中CO2浓度的垂直梯度和气体扩散系数求算土壤呼吸速度(于贵瑞和孙晓敏,2017)。箱式法采用不同类型的箱子罩住一定面积的土壤及其植被上方,隔绝箱子内外的气体自由交换,通过测定箱内的CO2等气体浓度变化来计算植被—大气或土壤—大气间的气体交换量。箱式法主要包括:静态气室-碱吸收法、静态箱-气相色谱法、动态(静态)气室-红外CO2分析仪法(于贵瑞等,2004b)。目前最为理想的方法是动态(静态)气室-红外CO2分析仪法,可以直接测定生态系统碳通量,并可以用气体采样法进行箱内多种气体的精细分析。然而,经过80多年的使用和发展,同化箱法始终存在改变被箱子罩住植被的生活环境即“箱体效应”的缺点,这也让测定结果很难反映实际的植被气体、水分交换情况(Kucera and Kirkham,1971)。
从20世纪50年代末开始,微气象原理被用于测定不同类型植被与大气间CO2和水汽交换,其主要通过观测大气湍流运动状况及待观测物质浓度来计算物质的通量值(Lemon,1960)。常用的观测方法有:空气动力学法(aerodynamics method)、热平衡法(heat balance method)及涡度相关技术(eddy covariance method)(Baldocchi et al,1988)。
空气动力学法又称为梯度法,基本假定是:在近地层中能量或物质的输送与其物理属性的梯度成正比,其比例系数(即湍流扩散系数K)受大气层结条件、气流垂直切变等影响湍流的外因参数的制约(于贵瑞和孙晓敏,2017),并用一个半经验的方程(1)来表示,将K分别代入到H2O /CO2扩散方程并积分得到H2O / CO2通量计算公式(2)、(3)。通过测定生态系统上部两个或两个以上高度的风速、气温、湿度、H2O和CO2浓度梯度,依据相似性理论间接计算H2O / CO2通量:
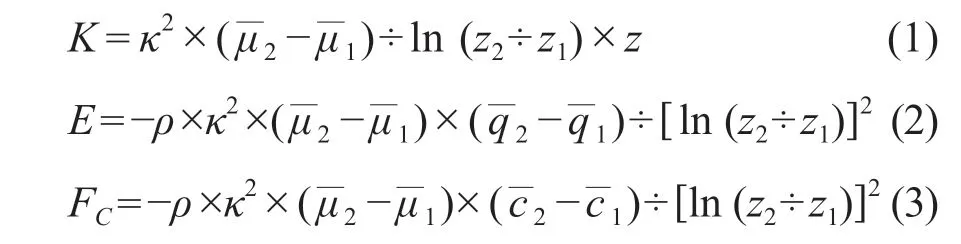
式中:κ为卡门常数,一般在0.35 — 0.43,通常取0.4;为风速切变;q1、q2分别为高度z1和z2上的比湿;ρ为空气密度;c1、c2分别为高度z1和z2上的CO2浓度观测值;E和FC分别为水汽通量和CO2通量。基于空气动力学原理的梯度法是利用两个或两个以上高度的观测数据求算通量的方法,而将两个高度中的一个高度确定为地面的通量计算方法称为整体法(于贵瑞和孙晓敏,2017)。热平衡法又称为热收支法,是利用梯度的观测资料计算H2O通量的方法之一,假定显热的湍流扩散系数与其物理量的湍流扩散系数相同,生态系统热量输入与输出平衡(沈艳等,2005)。之后在热量平衡的原理上,基于热量平衡方程与相似理论,根据波文比的定义提出了波文比-热量平衡方法,简称波文比法(关德新等,2004),根据彭曼公式提出了彭曼法(王健等,2002)。
所以空气动力学法和热平衡法都是通过物理量的垂直梯度来间接推算CO2/ H2O通量。涡度相关法与空气动力学法和热平衡法均不同,它是通过测定地表指定高度物理量与风速脉动的协方差来推算CO2/ H2O湍流通量的直接方法(张雷明等,2019),这种方法需要高精度,响应速度极快的湍流脉动测定装置。随着科学认知与观测技术的进步,20世纪70年代末80年代初,超声风速计和高性能的气体分析仪得以开发和改进,涡度相关技术逐步成为全球范围内连续CO2/ H2O通量研究的关键技术,并在20世纪90年代正式开展全球长期CO2/ H2O通量观测(Baldocchi et al,1996)。
作为涡度相关技术理论基础的雷诺平均和分解最早由雷诺(Osborne Reynolds)在1895年提出,后来随着流体力学和微气象学理论的长期发展,以及微气象学仪器、计算机和数据采集器等技术的进步,涡度相关技术逐渐成熟。涡度相关法最早在1951年被Swinbank应用于草地显热和潜热通量测定,开创了涡度相关法的应用先例(耿绍波等,2010)。20世纪70年代前后涡度相关技术已开始应用于不同类型生态系统通量的观测,20世纪80年代开始用于测定大气 — 群落间的CO2通量,至20世纪90年代,随着超声风速计和红外CO2/ H2O气体分析仪的研发和正式投入使用,涡度相关技术逐渐趋于成熟并开始被广泛用于CO2/ H2O通量的长期观测(于贵瑞和孙晓敏,2017)。20世纪90年代之前,传感器性能和数据采集系统的局限性一直限制着涡度相关技术在野外观测中的应用。当时只能在生长季进行短期测定,分析生态系统CO2通量的短期变化特征及其生物环境控制机制等问题(Verma et al,1986)。观测性能稳定、时间常数较短的商用红外气体分析仪的出现,帮助人们实现了用涡度相关技术测定从日尺度、月尺度到年尺度的生态系统CO2/ H2O通量。目前,应用涡度相关法测定CO2/ H2O通量时所采用的CO2分析仪器主要用两种类型:一种是开路红外气体分析仪,另一种是闭路红外线气体分析仪。开路与闭路系统的特点如表1所示,各种通量观测方法的优缺点见表2。
1997年,随着涡度相关技术的不断成熟,通量观测研究的区域性网络开始形成,欧洲通量网(CarboEurope)和美国通量网(AmeriFlux)相继建立,成为国际通量观测网络(FLUXNET)的基础。通过对陆地生态系统气候和生态系统代表性站点观测,国际通量观测网络正在提供背景值并直接测量生态系统生理生态过程如何对环境和生物强迫做出响应,以及在一个全球变暖、CO2增加的条件下生态系统生理生态过程如何变化(Baldocchi,2020)。2001年,我国正式启动建设陆地生态系统通量观测研究网络,2003年正式实现8个观测站点CO2/ H2O通量的联合观测研究,2014年,通过联合国内行业部门及高等院校观测站点,共同组建了我国通量观测研究联盟(ChinaFLUX),涵盖了我国主要农田、森林、草地等生态系统(于贵瑞等,2014)。目前涡度相关技术已成为国际通量观测网络和我国通量观测研究网络的主要技术手段(张雷明等,2019)。
1.2 模型模拟
涡度相关通量测定的优势在于确定生态系统碳通量的连续变化,认识碳循环生理生态过程机理及其与环境条件的关系。尽管现在已形成区域性和全球性的网络,但它仍然是一种小尺度观测方法,其结果本身只代表特定生态系统在特定环境中的碳循环特征,不能直接外推到区域尺度(曹明奎等,2004)。基于生理生态过程的模型模拟生态系统中能量、碳水循环是分析陆地生态系统碳水循环过程机制与研究生态系统与气候之间相互作用的一种有效手段(卢晓菲,2018),是定量表达CO2/ H2O通量时空特征,分析其内在机制的有效方法(刘晓文等,2020)。
关于陆地生态系统碳循环与碳交换量的评价模型研究,自20世纪50年代Craig(1957)在全球碳循环中用两个储库来模拟陆地生态系统的碳平衡以来,得到快速发展(于贵瑞和孙晓敏,2017)。最初是以统计模型或称为经验模型的CO2/ H2O通量研究模型为主,代表模型为Miami模型(Lieth and Whittaker,1975)、Thornthwaite模型(Uchijima and Seino,1985)。这类模型是将气候作为生态系统的主要控制因素,通过与气象资料如气温和降水等因子建立回归关系估算生态系统生产力。由于其将生态系统内在机理描述得较为模糊,并且只能以年为时间尺度,估算误差较大,难以准确描述CO2/ H2O通量的时空变化(刘晓文等,2020)。20世纪70年代以来,随着光能利用率原理的发现(Monteith,1972),光能利用率模型应运而生,代表模型有CASA模型(Potter et al,1993)、VPM模型(Xiao et al,2004)、GLOPEM-CEVSA模型(王军邦等,2009)等。该类模型建立在植物光合作用过程和光能利用率的概念上,认为任何对植物生长起限制作用的资源(如水、氮、光照等)均可用于NPP的估算(张美玲等,2014)。光合有效辐射是植被NPP的一个决定性因子,而植物吸收的光合有效辐射(APAR)则尤为重要。随着遥感技术的发展,APAR已可以通过遥感信息进行估算,这极大地改善了人们对生态系统CO2/ H2O通量研究的认知。然而,光能利用率模型不能从生理生态学机理上解释生产力的变化机制,尤其该模型在利用遥感提取的植被指数来模拟植被NPP的某些环节仍然存在一些不确定性和不一致性(朱文泉等,2005)。

表1 开路和闭路红外气体分析仪比较(于贵瑞和孙晓敏,2017)Tab.1 Comparison of open and closed circuit infrared gas analyzers (Yu G R and Sun X M, 2017)

(续表2 Continued Tab.2)
陆地生态系统在结构上包括细胞、组织、器官、植物个体、生态系统以及景观等多个层次,上层发生的各种生态行为基于底层所发生的各种生物化学及物理过程(巴特尔·巴克等,2008)。机理模型是指在模型中应用了生态系统层次性过程相互作用的生态学原理,通过模拟叶片尺度上的CO2、水分和能量交换的生理和物理过程(如光合作用、呼吸作用、蒸发蒸腾作用、分解作用和营养循环等过程)来进一步估算生态系统、景观等尺度上植被生长动态等的一类模型(康满春等,2019)。20世纪80年代,随着红外气体分析技术的发展,叶片水平CO2和水汽交换及其环境因子响应的直接测量方法取得了极大进步,推动了光合作用机理模型的发展(于贵瑞和孙晓敏,2017)。
Farquhar et al(1980)基于植被光合作用机理提出了光合作用生化模型(FvCB),认为叶片光合能力主要取决于光合羧化酶的羧化和电子传递两个基本过程。进入20世纪90年代中期,科学家们基于FvCB模型发展了多种生态过程机理模型,生态过程机理模型以生物物理模型和生物化学模型为代表(刘晓文等,2020)。生物物理模型考虑不同气象因子(气温、风速等)下植被与外界热量、动量和水的交换过程,并且综合考虑了不同气象背景下植被冠层对不同波段光谱的反射、吸收和散射等复杂过程及植被冠层蒸腾蒸发、水热转化等过程(谢馨瑶等,2018),能够揭示植被与气象因素的密切关系,代表模型有BATS模型、SIB模型(Sellers et al,1992)等。生物化学模型综合考虑气候、土壤植被条件等干扰因素,侧重于分析环境因子(气温、降水、大气CO2浓度等)变化对植被(叶、茎和根)CO2/ H2O交换及生态系统CO2/ H2O和养分循环的影响(任小丽等,2012),通过模拟生态系统的光合过程和蒸散过程等,估算土壤、植被与大气之间碳水循环变化。此类模型能够在全球气候变化背景下对全球和区域碳、水分和氮的循环过程、蓄积量及其分布格局等变化进行有效预测和评价,是预测和分析大尺度陆地生态系统在未来气候变化背景下物质循环和能量流动变化的有效工具(周洁等,2013),其代表模型有CENTURY模型(Parton et al,1993)、TEM模型(McGuire et al,1993)和CEVSA模型(Cao and Woodward,1998)等。由于生态过程机理模型反映的是植被和土壤类型不变且生态系统在平衡状态下的区域CO2/ H2O通量变化情况,不能揭示各个时刻植被结构和组成特征,因而出现了动态全球植被模型(dynamic global vegetation model,DGVM)(于贵瑞和孙晓敏,2017)。该类模型耦合了生物地球化学模型和生物地球物理模型,可以同时模拟陆地表面物理过程、冠层生理过程、植被动态过程、碳循环等过程,还能模拟全球气候变化和人类扰动(如土地利用/土地覆盖变化)等各种情境下植被的动态响应和植被功能性变化,因此适用于研究大尺度上陆地生态系统对环境变化的响应和反馈(于贵瑞和孙晓敏,2017),其代表模型为LPJ-TEM模型(Pan et al,2004)和DELM模型(田汉勤等,2010)。准确评估未来陆地生态系统碳收支需要结合植被变化和植被变化对气候和大气CO2浓度的反馈,因而未来碳水循环模型可能会向基于动态植被的机理模型方向发展(毛留喜等,2006)。穆少杰等(2013)在总结了朱文泉等(2005)陆地植被净初级生产力计算模型研究进展的基础上,给出了不同类型陆地生态系统CO2/ H2O通量模型的比较,发现具有物理基础的生态生理过程模型在模拟、预测净生态系统交换时具有较高的准确性与可靠性。
通过对现有陆地生态系统碳循环模型的综合分析,陆地生态系统碳循环模型的研究重点是对生态系统碳库内部循环过程以及生态系统碳库与大气圈碳库相互作用的精细描述。观测站的结果需要向大、中区域尺度进行扩展,其中最主要的方法就是结合区域尺度陆地碳循环模型,在空间化的植被和环境要素空间数据库支持下,进行尺度扩展和区域以及全球尺度碳收支的精确评价(于贵瑞等,2004b),并模拟未来气候变化和人类活动对碳循环过程的影响。生态系统碳通量观测建立了生态系统GPP和NEP模型知识数据库,为模型建立提供假设与验证、机理与关系的认识、模型参数化和模型模拟或预测结果检验的数据支持。因此需要综合应用观测和模拟的手段,全面揭示陆地生态系统碳水交换过程及变化机理(刘晓文等,2020)。
2 陆地生态系统碳汇能力和水汽通量的动态变化
2.1 碳汇能力的动态变化
自2020年9月以来,我国多次向国际社会宣布,将采取更加有力的碳排放政策和措施,预计2030年森林蓄积量将比2005年增加60亿m3。目前,越来越多的国家正在将碳中和目标转化为国家发展战略,提出无碳未来的愿景(陈雅如和赵金成,2021)。在无碳愿景中,陆地生态系统碳汇功能在调节大气CO2浓度和减缓全球变暖方面发挥着无法替代的作用(Yu et al,2016)。对典型生态系统的长期碳通量观测是准确定量评价陆地生态系统碳汇功能,分析生态系统长期碳通量动态变化特征的重要研究方法。
陆地生态系统类型复杂多样,如森林生态系统、草地生态系统、农田生态系统以及荒漠生态系统等,由于下垫面的复杂性以及人类活动的影响,不同陆地生态系统的碳通量变化和碳汇功能存在一定程度的差异。即使是同种生态系统其碳汇能力变化的时空差异也十分明显。本文分别从森林、草地、农田以及荒漠生态系统碳汇能力观测展开研究,进一步分析不同下垫面条件下碳汇能力变化规律。
2.1.1 森林生态系统
森林是陆地生态系统的主体,能量的再分配对区域乃至全球气候有着重要影响,森林生态系统是陆地生态系统中面积最大、最重要的自然生态系统,具有降低大气温室气体浓度、涵养水源、调节气候、维持生态平衡等重要功能(卢晓菲,2018)。《巴黎协定》第5条明确提出森林是气候解决方案的一个重要组成部分,通过森林生态系统的保护、修复和可持续管理来减缓气候变化(陈雅如和赵金成,2021)。所有森林生态系统,热带森林、温带森林以及寒带森林,均表现为碳汇(Rebane et al,2020)。
国内外学者针对森林生态系统展开了大量研究。国外研究起步较早,在20世纪60年代末和70年代初,人们开始利用梯度法在森林生态系统开展CO2通量的观测研究,但是对梯度法在高大森林植被上的适用性存在疑问。至20世纪90年代,人们对森林生态系统CO2/ H2O通量展开长期观测(于贵瑞和孙晓敏,2017)。Wofsy et al(1993)首次利用涡度相关技术测定了马萨诸塞州中部落叶林生态系统年尺度CO2通量,之后基于涡度相关技术的CO2/ H2O通量的测定在全球范围内得到了广泛应用。
国内基于涡度相关技术对森林生态系统碳通量的研究起步较晚。但自2002年成立我国第一批通量观测站以来,就把森林生态系统作为研究重点,我国通量网第一批8个观测站中有4个观测站为森林生态系统观测站。经过长时间通量观测,积累了大量宝贵的原始数据。Yu et al(2016)在总结了我国通量网22个森林通量观测站超过10 a以上的观测数据之后,对区域陆地生态系统碳收支进行了评估:截至2014年,我国森林生态系统NEP最高出现在中亚热带森林,为(550 ±258) g · m−2· a−1(以碳计)(平均值±标准差),其次是暖温带森林和北亚热带森林,分别为(492 ± 37) g · m−2· a−1和343 g · m−2· a−1。与暖温带和温带森林相比,冷温带森林表现出较低的碳汇。传统上,人们认为幼林和正在恢复的森林生态系统碳汇作用明显,大约100 a以上的森林在总碳吸收和生态系统呼吸之间达到平衡,是不重要的碳汇。然而,随着长期涡度协方差观测技术的广泛应用,发现古老森林仍可作为重要的碳汇。我国东北温带长白山森林生态系统的主要植被是生长超过200 a的阔叶混交林,其固碳能力为(308 ±116) g · m−2· a−1(以 碳 计),表 现 为 强 大 的 碳 汇(Zhang et al,2006)。我国东南地区亚热带原始森林生态系统固碳能力在230 — 489 g · m−2· a−1,平均固碳能力为390 g · m−2· a−1(以碳计),同样表现为强大的碳汇(Yan et al,2013)。我国西南热带雨林生态系统的固碳能力在98 — 235 g · m−2· a−1,平均固碳能力为168 g · m−2· a−1(以碳计),也表现为碳汇(Zhang et al,2010)。相比原始森林,我国在西北、华北和东北地区展开了广泛的植树造林和退耕还林工程,根据第九次全国森林资源清查(2014 — 2018年)结果,人工林面积占全国林地森林面积的36.45%,人工林蓄积占全国的19.86%,相比第八次森清,森林蓄积人工林上升超过2%(崔海鸥和刘珉,2020),人工林的碳通量研究也越来越重要。我国东北温带地区的落叶松人工林在2004年5月 — 2005年4月表现为碳汇,其碳汇能力为146 g · m−2· a−1(以碳计)(Wang et al,2008)。在崔崧(2007)研究中,落叶松人工林的固碳能力年际变化很大,不同年份碳通量可以相差近一倍,但均表现为明显碳汇。总体来看,人工林碳汇能力弱于天然林,但是也要多方面考虑气候变化等多种方面的影响。
我国各气候带森林生态系统碳通量变化表现出明显的季节动态和年际差异,在我国东北长白山森林生态系统中,夏季90 d的时间贡献了全年GPP的67.0%和生态系统呼吸(RE)的58.9%,GPP和RE的季节变化趋势与温度密切相关(Zhang et al,2006)。我国华北地区天然落叶阔叶林生态系统中,NEE在生长季(5 — 10月)日尺度上,变化呈“U”字形;在季节尺度上则呈单峰曲线变化规律,全年NEE表现为明显碳汇(李润东等,2020)。在我国西南西双版纳森林生态系统(2003 — 2008年)的涡度通量监测中,生态系统总碳吸收(GEE)和RE之间有着相似的季节变化规律:在雨季增加而旱季减少,只是在雨季RE增加超过了GEE,在雨季表现为净碳源,而在旱季相反(Zhang et al,2010)。上述研究表明:对森林生态系统的研究,目前主要在于明确森林生态系统碳循环过程和碳汇能力,研究森林生态系统碳通量季节变化及长期变化的生物和环境控制机理。
2.1.2 草地生态系统
草地生态系统是陆地生态系统的重要组成部分,是我国陆地最大的生态系统。应该充分发挥草地生态系统对CO2等温室气体的吸收、存储、替代和适应功能。由于草地生态系统植被高度低、冠层结构简单、下垫面性质均一,常用箱式法和涡动相关法对草地生态系统CO2/ H2O通量进行长期观测,主要从日、季节、年际等不同时间尺度对典型草地类型进行研究(李琪等,2010)。一般认为,草地生态系统的碳固定大于碳排放,表现为碳汇功能(沈晓坤等,2014)。
箱式法在草地生态系统中的研究多以季节或年际为时间尺度,一般对草地生长季节进行定时间间隔取样分析。孙殿超等(2015)利用箱式法对科尔沁围封和放牧两种管理类型的沙质草地进行一个生长季的观测研究,发现围封17 a样地的生态系统生产力(GEP)、RE、蒸散量(ET)均最大,超过了围封22 a样地以及放牧样地,草地CO2/ H2O通量之间存在显著的线性正相关关系,认为围封能够使退化沙质草地生态系统的碳水循环速率提高,但围封时间不宜过久。王祥等(2017)利用箱式法对新疆昭苏盆地围封3 a的人工草地和天然草地CO2/ H2O通量进行观测,发现天然草地年平均NEE小于人工草地,全年平均ET却大于人工草地;天然草地CO2/ H2O通量之间存在极显著相关关系,而人工草地相关性不显著;天然草地翻耕重建成人工草地,草地生态系统NEE提高的同时ET降低,提高了水分利用效率和草地生产力。
涡度相关技术在草地生态系统CO2/ H2O通量观测中应用十分广泛。国外学者运用涡度相关技术在草地生态系统的研究较早,由Swinbank于1951年首次提出并应用于草地研究,之后人们利用涡度相关技术针对草地生态系统CO2/ H2O通量进行连续观测,对不同时间尺度碳水动态展开研究。通过长期连续观测通量,发现草地碳汇明显低于森林,具有显著的区域差异(Yu et al,2013),温带草原和高山草甸表现为弱碳汇,分别为(24 ±83) g · m−2· a−1和(45 ± 59) g · m−2· a−1(以碳计)。相比林地,草地更容易受气候变化等因素干扰,我国北方草原受干扰之后可能表现表现为碳中性或弱碳源(Yu et al,2016)。在我国华北和西北地区的内蒙古草原,水分条件限制作用明显,其中在我国北方半干旱农牧交错带的科尔沁沙质草地,其受到水分条件的限制表现为弱碳源,生态系统NEE为91.39 g · m−2· a−1(以碳计)(牛亚毅等,2018)。
草地碳通量的季节变化和年际变化明显,非生长季草地生态系统碳通量日变化不明显,波动多且昼夜变化不明显,主要表现为碳源(李琪等,2010)。生长季草地生态系统日间CO2吸收量和夜间CO2排放量都开始增大,出现明显的CO2吸收和释放峰值。草地生态系统NEP日动态具有单峰型(徐世晓等,2005;赵亮等,2008)和双峰型(薛红喜等,2009)。草地类型及所处环境不同,导致植被的光合作用、土壤呼吸以及各种环境因子季节变化的差异,进而会影响到草地生态系统NEE的季节动态特征。通常而言,5 — 9月为我国草地生态系统的碳吸收期,以7月、8月碳吸收最强,同时碳释放最大值也往往出现在这一时期(毛留喜等,2006)。华北地区内蒙古草原生态系统的碳通量日变化表现出单吸收峰特征,而在生长季后期表现为双吸收峰。NEE日最大累积吸收量和最大累积释放量出现在植被生长最旺盛的7月、8月(薛红喜等,2009)。在青藏高原三江源人工草地生态系统中,日最大碳吸收量出现在7月,日最大碳吸收率和最大碳排放率都出现在8月(赵亮等,2008)。同林地相似,对草地生态系统的研究,主要明确其碳源/汇能力,研究草地生态系统碳通量季节变化以及年际变化规律,确定控制其变化的生物和非生物机理。同时,通过半干旱地区草地和荒漠生态系统的对比研究,明确草地与沙丘之间的差异,可以为减少草地荒漠化提供参考。
2.1.3 农田生态系统
农田生态系统是人类活动最频繁、最剧烈的生态系统。农田生态系统处于近地面层,群落结构单一,营养结构简单,生态系统稳定性差(李晶等,2003)。农田碳通量的差异很大,我国农业生产整体上表现为碳汇,且我国农业生产的碳排放量、碳汇量和净碳汇量三者均呈增加态势,碳汇增速明显快于碳排增速(田云和张俊飚,2013)。
涡度相关法是农田生态系统碳通量观测的主要方法,国内外学者利用涡度相关技术对农田生态系统碳通量进行了大量研究。由于农田空间差异性很大,不同地区农田通量差异巨大。位于我国温带半干旱气候区的农田生态系统碳通量明显小于全国其他地区,主要种植一些耐旱作物,如小麦、荞麦、玉米等,其碳汇能力在75 g · m−2· a−1(以碳计)左右(Yu et al,2016);在作物成熟后,农田会损失76%的生物量,这极大地降低了农田生态系统的固碳能力(Zhang et al,2007)。在我国温带季风气候区的玉米/小麦轮作生态系统中,年均碳汇能力约为559 g · m−2(以碳计),小麦和玉米的NEP季节总量分别为303 —395 g · m−2和201 — 244 g · m−2(以 碳 计)。小 麦碳汇能力强于玉米,但均表现为长期碳汇。作物收获时,麦田会变成弱碳汇而玉米田则变成了弱碳源(Lei and Yang,2010)。我国亚热带农田生态系统碳汇能力明显增强,其碳汇能力达到了2475.6 g · m−2· a−1(以碳计),是大气CO2显著的汇,这与其湿润的气候条件和科学的耕种方式有很大关系(朱咏莉等,2007)。在我国由半干旱气候区向温带气候区和亚热带气候区的过渡过程中,主要耕作物发生了改变,农田碳通量也在明显增加。
20世纪80年代,我国已在农田生态系统水热运动的实验研究中取得了较大的成果。但由于农作物的生育周期较短,农田的CO2通量研究通常以季节性或日尺度为主(马虹等,2012)。我国西北干旱区绿洲玉米生态系统生长季不同阶段的碳通量日变化差异很大,在天亮后和黄昏前均表现碳吸收,且在12∶00 — 14∶00碳吸收能力有轻微下降(Ji et al,2011)。梁涛等(2012)对我国东北温带气候区玉米农田生态系统生长季(5 —10月)及非生长季CO2通量动态变化进行了分析,发现非生长季NEE日动态趋势不明显,生长季日动态呈明显的“U”型分布,全年碳收支总体表现为碳汇。孙小祥等(2015)对我国长三角地区典型稻麦轮作农田生态系统的CO2通量进行分析,发现稻麦轮作生态系统NEE具有明显的日变化和季节变化特征,总体是明显的碳汇。NEE月平均日变化总体呈“U”型曲线;NEE季节变化则呈显著的“W”型双峰特征,分别对应小麦、水稻的生长季。吴东星等(2018)对华北地区冬小麦农田生态系统CO2通量特征进行分析,发现农田生态系统NEE日变化过程在生长季呈单峰“U”型、双峰型或不规则型曲线。在作物不同生育期,农田生态系统NEE也有明显差异,一般来说,作物在营养生长期固碳能力大于生长初期或生殖生长期,碳固定和排放能力往往在农作物生长最旺盛的时期同时达到最大(薛红喜等,2012)。农田生态系统碳通量特征受农作物种类、区域水热条件、种植方式、作物生长阶段的控制(郭家选等,2006;李俊等,2006;李琪等,2009)。
2.1.4 荒漠生态系统
荒漠生态系统约占全球陆地面积的35%,我国干旱和半干旱地区约占陆地国土面积的40%,是对CO2浓度升高响应敏感的生态系统之一。干旱荒漠生态系统较单一的植物物种组成和较低的植被覆盖率,使其抵御外界干扰的功能明显弱于其他生态系统,因而荒漠生态系统能很快地表现出对环境变化的响应与反馈(刘冉等,2011)。有学者对荒漠生态系统的碳吸收进行研究,将荒漠区作为大气CO2“未知碳汇”的分布区以及未来隔离化石燃料CO2排放的潜在重要区进行研究(周琪等,2014)。我国西北的部分荒漠区为弱碳源区(刘冉等,2011)。
与森林、草地以及农田等生态系统相比,利用涡度相关技术对干旱、半干旱荒漠地区CO2/ H2O通量的连续观测研究较少(刘冉等,2011)。在我国,荒漠主要集中在西北内陆地区,为温带大陆性气候。荒漠生态系统主要以梭梭群落为主,并伴有一年生草本,在荒漠与草原过渡地区,其主要优势种为半灌木、灌木和草本。我国西北新疆准格尔盆地沙漠地区,荒漠生态系统在旱季表现 为 弱 碳 汇 或 碳 源((5 ± 12) g · m−2· a−1),但 是在雨季表现为较强的碳汇((40 ± 12) g · m−2· a−1)(以碳计),碳通量差异与地区降水有着明显关系(Liu et al,2012)。在草原化荒漠和荒漠化草原过渡地区,植被恢复是治理沙漠化的重要手段,植被恢复生态系统在集中大部分全年降雨时的生长季表现为碳汇,在有效降雨之后的1—2 d,荒漠生态系统可以实现碳源向碳汇的转变。虽然相比湿润地区荒漠生态系统碳汇功能较弱,但是由于我国西北荒漠地区面积巨大,使得荒漠生态系统存在巨大的碳固定潜力(Gao et al,2012)。我国北方半干旱区荒漠生态系统表现为碳汇,其中榆林站和通辽站观测值分别为132.8 g · m−2· a−1和59.6 g · m−2· a−1(以碳计),同样的荒漠地区其碳通量差异仍十分显著(龚婷婷等,2018)。
在荒漠生态系统的季节变化和年际变化中,天山荒漠生态系统虽以地下水而不是降水为主要补给水源,但是CO2通量并没有同地下水一样表现出季节、年际的稳定特征,而是随着降水时间和强度分配不均表现出明显差异,主要原因是降水差异导致生态系统一年生草本植物在群落中占比不同,进而影响了群落的初级生产力(刘冉等,2011)。我国北方荒漠区在进入生长季时,随着光合作用增强,GPP和RE逐渐增强,同时生态系统NEP逐渐增大,进入生长旺盛阶段,各碳通量均达到峰值,在生长季后期进入非生长季,碳通量逐渐减少并在0附近波动(龚婷婷等,2018)。综上所述,对于荒漠生态系统而言,国内研究运用涡度相关技术进行大尺度、长序列观测的研究还在初步阶段,主要研究水分条件对碳通量的影响,这主要是因为荒漠区降水稀少,植被长期处于水分亏缺状况,碳通量对于水分的变化较敏感(龚婷婷等,2018)。
2.2 水汽通量动态变化
生态系统H2O通量的观测主要包括植被蒸腾、冠层截留蒸发以及土壤蒸发三部分通量之和(于贵瑞和孙晓敏,2017)。H2O通量可以通过涡度相关系统进行直接观测,一般将H2O通量定义为单位时间内通过单位面积所输送的水汽量。ET在涡度相关系统能量闭合的基础上通过潜热通量进行计算。通常使用的蒸散强度就是H2O通量,其时间上的积分值称为ET。ET和H2O通量是重要的水分状况指标。ET是陆地生态系统水循环和能量平衡的重要组成部分,研究ET的时空变化及其影响因素对水资源管理和评估具有重要意义(Nemani and Running,1989)。涡度协方差技术通过直接测量大气和生物圈之间的H2O通量,在探索ET的时空变化及其影响因素中发挥着重要作用(Yu et al,2016)。通过涡度通量网的长时间观测,我国通量网分析了典型生态系统中H2O通量日变化、季节变化和年际变化过程,理解控制水汽通量的环境因素和生物因素,并揭露ET的空间变化特征(Yu et al,2016)。2003 — 2006年我国西南亚热带雨林年均ET占降水量的78%,旱季11月 — 次年2月日均ET为2.6 mm,雨季5 — 10月日均ET为3.1 mm,旱季ET明显高于降水量(Li et al,2010)。我国东南地区亚热带针叶林日均ET为5.34 mm,在经历强干旱时日均ET下降到1.84 mm,之后恢复到4.8 mm(Tang et al,2014);同时,2003 — 2010年我国东南亚热带针叶林年际ET平均值为786.9 mm,标准差103.4 mm(变异系数为 13.1%)(Xu et al,2014)。我国东北暖温带季风气候区芦苇沼泽的年均ET为452 mm,在生长季最旺盛的时段,日均ET达到3.6 mm,生长季日均ET为2.32 mm,非生长季的ET贡献了全年ET 的 13% — 16%(Zhou et al,2010)。Gu et al(2008)对我国青藏高原高山气候下嵩草草甸2002 — 2004年ET进行了研究,观测期间其年ET均值为391 mm,占年降水量的60%左右,生长季日均ET为2.12 mm,非生长季日均ET很低,小于0.4 mm。在不同气候带和不同植被类型ET整体量化分析中,我国整体ET为(534.7 ±232.8) mm · a−1。基于植被类型分析,阔叶森林的ET最高,其次为农田,且灌溉农田明显高于非灌溉农田,最低为草原和荒漠生态系统。在不同气候带中,热带季风和亚热带季风气候区ET最高,其次为高山性气候区和温带大陆性气候区,ET分布表现出与降水分布相似的规律(Zheng et al,2016)。
在ET日变化、季节变化和年际变化过程中,不同生态系统同样表现出明显差异。在西双版纳热带雨林生态系统中,无论旱季或雨季,ET日变化均呈“单峰型”。旱季,中午净辐射达到最大值后的2 h左右,ET达到最大值。雨季ET日变化基本遵循净辐射和感热通量变化规律。在季节变化过程中,从11月到次年1月ET呈下降趋势,2月份开始ET略有增加,并保持增加趋势。但由于旱季上层土壤含水量较小,导致整个旱季生态系统ET没有显著变化。在5 — 10月的雨季,生态系统ET值较高,变化较大。同时,ET的年际变化同样显著,旱季水分蒸发量的主要来源是前一个雨季储存的土壤水分,不同年份储存的土壤水分的差异,是造成ET年际差异的重要原因(Li et al,2010)。我国东南地区亚热带针叶林经常会发生季节性干旱,在发生干旱的年份,生长早期(1 — 182 d)ET会持续增加,在发生干旱时ET明显下降,并在经历干旱之后逐渐恢复,在生长后期(238—265 d)由于植物生理活性下降和大气条件的改变,ET会再次下降(Tang et al,2014)。在对我国东北暖温带季风气候区芦苇沼泽的研究中,发现ET年际差异主要体现在6 —9月的生长季,在生长季ET可能呈现单峰或双峰变化(Tang et al,2014)。高山草甸生态系统ET日变化表现出“单峰型”,夜间ET值较低且变化较小,白天迅速增加,12∶00 — 14∶00达到峰值,之后开始下降,18∶00 — 20∶00接近0,并且发现ET对不同气象因子的响应具有滞后性(Zheng et al,2014)。
2.3 碳通量与水汽通量的关联
碳通量和H2O通量作为生态系统功能的关键,CO2/ H2O通量之间的联系至关重要。针对CO2/ H2O通量耦合变化,水分利用率(WUE)是连接生态系统碳循环和水循环的重要物理参数。长时间序列涡度观测CO2/ H2O通量数据为WUE的研究分析奠定了基础。由于具体研究的时空尺度不同,WUE的定义方式也多种多样,最基本的定义是CO2通量和H2O通量的比值。因此,在生态系统水平和冠层水平的WUE可以定义为净生态系统生产力与蒸散量的比值(NEP/ET)或者净初级生产力与蒸散量的比值(NEE/ET)。在涡度系统的研究中,主要分析GPP/ET的变化(Kang and Cho,2021)。典型生态系统中,森林生态系统年均WUE从温带阔叶林的(9.43 ± 1.28) mg · g−1,到亚热带针叶林的(9.27±1.77) mg· g−1和亚热带阔叶林的(6.90 ± 0.69) mg · g−1(以CO2、H2O质 量 计),从温带到亚热带的跨越,森林生态系统GPP和ET之间存在不同的关系(Yu et al,2008)。在草原生态系统中,高山草甸和华北温带草原具有广泛的代表性,高山草甸生态系统年均WUE为0.79 mg· g−1,温 带 草 原 生 长 季 平 均WUE为0.80 mg· g−1(以CO2、H2O质量计)。不同位置的高山草甸同样存在水分限制,在相对干旱的草地生态系统中,无论是高山草甸还是温带草原,其WUE的年际变化与GPP变化一致,这表明GPP的年际变化大于相对应的ET。而且对比分析发现高生产力的生态系统同样拥有高WUE的空间分布格局(Hu et al,2008)。农田生态系统中,我国华北平原种植大面积的小麦,随着小麦生长阶段的不同,WUE呈现出明显的阶段性变化。返青至拔节期间,小麦平均WUE为2.76 mg · g−1,拔节至抽穗期间平均WUE为6.27 mg · g−1,之后至小麦成熟平均WUE为5.39 mg · g−1(以CO2、H2O质量计)。在小麦的生长旺盛期WUE明显增加,随小麦逐渐成熟和叶片衰老而逐渐降低。与GPP明显的季节变化不同,WUE的季节变化不太显著,主要是组成WUE的GPP和ET季节变化线性关系密切(Zhao et al,2007)。同样地,作为主要粮食作物的水稻在我国广泛种植,东北地区水稻年平均WUE为1.00 mg · g−1,生长季平均WUE为1.35 mg · g−1(以CO2、H2O质量计)。水稻WUE同样表现出阶段性变化,生长季刚开始WUE通常较小,变化相对较稳定,在之后几周,WUE迅速增长,剧烈波动并达到最大值,最高值达到4.1 mg · g−1(以CO2、H2O质量计),之后开始下降直到生长季结束(Wang et al,2017)。在不同生态系统中,森林生态系统的WUE最高,这也体现出了森林生态系统强大的碳吸收能力。
3 碳水通量影响因子分析
3.1 碳通量影响因子
在不同时空尺度下,碳通量反馈过程对各种气象、水文和生态因素都很敏感(Ge et al,2011)。目前已经有大量的研究分析主要环境胁迫因素和气候变化对陆地大气碳通量的影响(Schmidt et al,2011;Wagle et al,2015)。通过模型模拟以及时间序列分析,生态系统碳通量对气候变化的响应可以划分为气候效应和生物效应(Shao et al,2014)。在短时间尺度(如每日)碳通量变化很大程度上是由气候效应控制,例如太阳辐射、温度、降水等因素(王海波等,2014);在较长时间尺度(如季节性和年际),碳通量变化是由生物对气候变化的反应控制(Shao et al,2014)。
温度、水分、光合有效辐射、CO2浓度等是导致不同生态系统碳通量变化的主要环境因素,生态系统碳通量对各要素的敏感性有所不同。温度包括空气温度和土壤温度,是控制生态系统碳通量的重要因子,对于森林生态系统而言,温度的升高一方面影响叶片气孔导度,影响森林植被的蒸腾作用和光合作用,降水条件不变的情况下仍可能产生水分胁迫,对森林碳吸收功能产生影响;另一方面,增温导致呼吸速率加快,降低森林生态系统碳汇功能(宋春林等,2015)。Chan et al(2018)研究造林14 a后树林的碳通量变化,发现土壤温度分别解释了23%、43%的GEP和RE变化。李润东等(2020)对温带落叶阔叶林碳通量的研究表明土壤温度是影响夜间NEE的主导因子。草地生态系统中不同的草地类型对温度响应有明显差异,青藏高原东缘高寒草甸生态系统夜间生态系统RE与土壤温度具有较好的指数相关关系,随着昼夜温差增大,CO2净吸收量增加(王海波等,2014);而西藏高原草原化嵩草草甸生态系统随着昼夜温差增大,CO2净吸收量反而减少(石培礼等,2006)。天然草地NEE对温度的响应显著高于人工草地,天然草地的温度响应更有利于有机碳的分解,而人工草地则更有利于碳的固定(王祥等,2017)。农田生态系统中土壤温度是控制碳通量的重要因子,稻田夜间生态系统呼吸速率与土壤温度呈指数相关,5 cm处土层温度是反映稻田呼吸速率变化的最适宜温度指标(朱咏莉等,2008)。小麦各生长阶段对温度的响应不一致,冬小麦分蘖期和灌浆期NEE日总量与土壤温度呈显著的相关关系,越冬期和拔节期的相关关系不显著。分蘖期、越冬期和灌浆期小麦NEE日总量与土壤温度呈正相关关系而拔节期则呈负相关(吴东星等,2018)。春夏两季玉米地日尺度NEE变化受温度影响存在差异,但均呈单峰曲线关系。夏玉米NEE随气温和土壤温度的升高而逐渐降低,春玉米NEE随土壤温度和气温的升高呈先上升后下降的趋势(陈宇,2019)。荒漠生态系统中温度变化会造成碳固定能力变化,在植被生长初期和后期,高温会导致该生态系统NEP增加,生长旺盛阶段,高温状况下荒漠植被的碳固定能力则表现出抑制作用(龚婷婷等,2018)。温度作为影响碳通量的重要环境因子,在植被不同生长阶段表现出不同的影响机制,并可以通过温度变化改变植物对水分等其余环境因子的需求来间接影响生态系统的碳通量变化。
水分条件包括降水和土壤含水量,降雨量和降雨时空变异性会直接影响生态系统中植被的生长,进而对生态系统固碳能力产生影响。在不同生态系统中,水分条件对固碳能力的影响表现出相似结果。降雨量可以改变土壤中的含水量,进而影响土层储水量,对生态系统碳通量日变化和季节变化产生影响(Machado et al,2016)。在土壤含水量较低的情况下,植被叶片功能受到限制,气孔关闭,导致光合能力下降,影响植被生产力,碳吸收量显著减少;当土壤水分过高时,会造成土壤通透能力减弱,影响根系呼吸,致使根系的吸水能力减弱,影响作物的碳汇能力。只有在适宜的土壤水分条件下,才能达到最大的光合效率,间接提高生态系统的碳汇能力(林同保等,2008)。需要注意的是,由于荒漠生态系统大多分布于干旱、半干旱气候区,受到水分有效性的制约,植被遭受更多的水分胁迫,使得荒漠生态系统中水分状况对NEE影响最大(张晴和李力,2009;刘冉等,2011;周琪等,2014)。水分条件作为重要的环境因子,时刻参与生态系统碳通量调节,过高或过低的水分条件均会抑制生态系统的固碳能力。只有在适宜的水分条件下植被光合作用才有机会达到最大,不受水分条件的限制,会间接提高生态系统的碳汇能力。
光合有效辐射是决定GPP、影响NEE变化的主要因素,太阳辐射是陆地生态系统CO2/ H2O通量的主要驱动因子,光照是植物生长的重要因子。当生长季温度适宜时,生态系统的初级生产强烈依赖于光照条件,光和有效辐射与生态系统NEE之间有明显的相关关系,随着光合有效辐射的增加,作物对碳的同化吸收能力加大。光合有效辐射是直接影响温带落叶阔叶林NEE的主导因子(李润东等,2020),人工混交林NEE随光合有效辐射的增大呈线性增加(同小娟等,2009)。高寒草甸日间生态系统NEE主要由光合有效辐射控制(石培礼等,2006;王海波等,2014;宋春林等,2015)。另外,生态系统碳通量存在光饱和现象,在碳交换日过程中,NEE并不完全随着光合有效辐射的增加而增大,当光合有效辐射超过某一值时,NEE随光合有效辐射的增加而降低(赵亮等,2008)。冬小麦农田生态系统不同生育期NEE与光合有效辐射的相关关系也达到极显著水平(吴东星等,2018)。光合有效辐射作为直接影响光合作用的环境因子,在碳通量变化过程中扮演着重要角色,温度适宜条件下光合有效辐射的增加在植被光合作用达到生理限制之前会显著提升生态系统的碳通量。
目前多数研究表明大气CO2浓度增加可对森林生态系统产生“施肥效应”,可以提高植被光合速率,进而提高生态系统固碳能力(Fernández-Martínez et al,2019)。由于森林生态系统是地球最大的碳库,因此CO2浓度对生态系统碳通量的影响也主要集中在森林生态系统中,CO2浓度的增加总会伴随NEE的增加,但受到全球气候变暖的抑 制(Fernández-Martínez et al,2019)。虽 然 目前研究表明CO2浓度增加会提高生态系统固碳能力,但其内在物理机制以及长期效应仍是未知的,这种模式最终可能会随着土地碳汇的饱和而发生逆转。
气候效应驱动碳通量的短期变化,而生物效应驱动在长时间尺度上可能更重要。生物效应对NEE年际变异的相对贡献随着时间尺度的增加而增加,与此同时,气候效应的相对贡献逐渐减少。生物效应主要是植物本身的光合作用与呼吸作用潜力,而反映光合与呼吸作用潜力的两个指标为最大光合速率与参考呼吸速率。年尺度生物效应对我国亚热带常绿人工林、美国落叶林和美国草地生态系统NEE的贡献分别为47%、69%和69%。研究还发现,在我国亚热带常绿人工林生态系统中,2003 — 2009年的年尺度气候和生物效应之间有很强的负相关,这表明这两种效应相互抵消,最终结果表现为相对稳定的碳汇(Shao et al,2014)。在千烟洲常绿林中,生物效应对NEE年际变异的贡献为47%,气候的变动除了直接影响碳通量,可能还会通过改变生态系统的营养条件与叶面积指数等特征进而对生物效应产生影响(Ge et al,2011)。
3.2 水汽通量影响因子分析
通过研究世界不同地区ET变化的影响因素,气候变化被认为是过去半个世纪影响ET变化的主要因素(Wagle et al,2016)。在不同地区,ET的空间变异性很大,常绿林ET高于落叶林、阔叶林ET大于针叶林、亚热带低纬度地区ET高于高纬度温带地区(路倩倩等,2015)。
不同森林类型的ET与土壤含水量的关系不尽相同,土壤含水量在受水分条件限制的森林生态系统中是控制生态系统蒸散的主要因素。而且,森林ET对降水的响应存在一个阈值,阈值前后两者相关关系从正相关转变为负相关(林友兴等,2019)。在干旱荒漠区生态系统的蒸散过程中,地下水水位的变化影响植被生长和覆盖度变化,进而对整个生态系统蒸散过程产生影响,因而采用地下水水位而不是土壤含水量定量表达荒漠生态系统的水分胁迫过程更加准确(袁国富等,2015)。农田生态系统中由于存在灌溉等人为因素的影响,使得农田生态系统中水浇地ET与降水量和灌溉量之和显著相关,而旱地ET则与降水量显著相关(莫兴国等,2005)。
净辐射是控制我国东部森林生态系统ET季节变异的最重要因素,且水分越充足,ET与净辐射之间的相关性越强(路倩倩等,2015),这一结论在北京人工林生态系统中也得到验证,林地ET与净辐射的正相关性在雨季较旱季更加显著(刘晨峰等,2009)。因此比较净辐射和水分条件的关系可以发现:净辐射是ET发生的条件,不同的水分条件决定了净辐射对ET的影响程度。同时,在对ET影响因子进行研究的过程中,人们发现ET对环境变量的响应存在滞后现象,ET日变化过程与净辐射相似,但与空气温度和饱和水汽压差存在明显的相位差,ET在相同气温或饱和水汽压差的情况下,上午明显大于下午,使得ET — 气温与ET — 饱和水汽压差关系曲线呈顺时针绳套。通过研究分析,净辐射和气温或饱和水汽压差之间的相位差的变化是调节ET — 气温或饱和水汽压差滞后强度的重要指标(Zheng et al,2014)。
在全球水通量站点的研究中,ET在区域和全球范围内有显著的年际变化,1982 — 2002年全球ET以15 mm · a−1的速率不断增加。湿润地区ET受太阳辐射的控制,而太阳辐射与云量和气溶胶的变化有关。干旱地区则还需考虑水分条件的限制(Wang et al,2010)。2000年之后,我国ET在15 a内整体呈轻微下降趋势并且具有年代差异,前10 a整体呈显著下降趋势,后5 a呈轻微上升趋势,其中净辐射是影响年际差异的主要因素(牛忠恩等,2019)。实际上,各气象变量之间存在较强的相关性,它们对生态系统CO2和水热通量的影响常常交织在一起(王秋凤等,2004),仅仅独立考察单个因子变化对生态系统的影响是不够的,在今后的研究中还应加强对气候变量综合影响的分析。
3.3 水分利用率影响因子分析
在CO2通量和H2O通量的动态变化中,存在着“耦合和解耦”的现象,在“耦合和解耦”现象的背后存在一系列环境和生物控制机制。涡度站对典型生态系统CO2/ H2O通量的长期连续观测为WUE的计算和分析提供了数据基础。在我国东部森林生态系统样带上,不同地带森林生态系统GPP和ET对气温、饱和水汽压差以及净辐射的响应差异很大。温带森林碳固定和水流失具有强烈的相关性,气候因子的季节变化对光合作用和蒸散的影响大致相同,从而使整个生长季WUE相对稳定并保持在较高水平;而在亚热带森林中,GPP和ET之间对气候因子的响应是非同步的,这也导致生长季WUE降低。控制整个东部森林生态系统WUE空间格局的主要因素是年平均气温和年平均降水,可以解释WUE变化的85%(Yu et al,2008)。在对我国草地生态系统样带的分析中发现,WUE随叶面积指数的增加而增加,而与气温和降水的增加无关,光合过程是WUE季节变化的主导调节因子(Hu et al,2008)。对生态系统站点的统一观测和分析表明:由于气候梯度影响生态系统类型的分布,使得WUE随海拔升高呈现下降趋势。随着年平均温度、年平均降水和平均叶面积指数的增加,WUE增加明显,但是年平均光合有效辐射的增加会抑制WUE。进一步分析发现:平均叶面积指数和海拔方程可以描述WUE空间变异的65%,年平均温度和年平均降水对WUE空间变异的影响可能是通过改变平均叶面积指数来实现,而年平均光合有效辐射的机制仍需进一步研究(Zhu et al,2015)。
4 存在问题与展望
目前陆地生态系统CO2/ H2O通量研究发展十分迅速,取得了丰富的研究成果。但在实际的研究中仍需要进一步完善,今后的研究工作还需要在以下几个方面进行加强:
(1)将涡度相关技术与模型模拟的方法进行有机结合,加强由观测站点向区域尺度以及全球尺度扩展的研究力度,将有助于进一步研究陆地生态系统碳水循环对全球气候变化的正负反馈机制。
(2)模拟模型在驱动数据、模型结构和模型参数等方面存在着很大的不确定性,未来研究中,应着力于降低驱动数据的系统误差和随机误差,深入理解生态系统关键过程和控制机制,构建完整的参数计算和获取体系。
(3)因经济成本和数据代表性问题,通量观测站多位于典型生态系统内部,较少关注生态系统过渡带的CO2/ H2O通量,未来应加强对各典型生态系统之间平缓过渡地区的CO2/ H2O通量观测研究。
(4)生态系统中CO2/ H2O通量的变化通常是各种环境因子和生物因子综合影响的结果,各因子之间的相互作用共同影响CO2/ H2O通量。目前对CO2/ H2O通量影响因子的研究主要集中在环境因子方面,主要关注CO2/ H2O通量与环境因子之间的相互关系,环境因子与生物因子复杂的共同作用对CO2/ H2O通量的影响及其反馈仍需要进一步展开研究。
随着涡度相关技术在陆地生态系统中持续测定时间的增加,区域或全球尺度的年际CO2/ H2O通量变化的研究越来越多,气候效应与生物效应对CO2/ H2O通量的影响研究也越来越深入。这要求涡度相关技术与模型模拟方法的有机结合,更加准确地预测全球CO2/ H2O通量变化。气候效应以及生物效应之间的相互作用,以及多因子对CO2/ H2O通量变化的综合影响,决定了CO2/ H2O通量与气候变化之间复杂的反馈机制,这也是未来碳水通量研究的一个主要方向。
猜你喜欢
河北地质(2022年2期)2022-08-22
农业灾害研究(2022年1期)2022-05-07
环境卫生工程(2021年1期)2021-03-19
科学大众(2020年17期)2020-10-27
农业灾害研究(2019年3期)2019-10-21
中国海洋大学学报(自然科学版)(2019年11期)2019-10-12
热带气象学报(2019年6期)2019-03-02
科技资讯(2018年30期)2018-03-11
现代园艺(2017年23期)2018-01-18
应用海洋学学报(2015年2期)2015-11-22
