
中国西部地区归化植物时空分布特征研究
2022-04-08 01:08李龙沁许光耀高越莫训强李洪远
广西植物 2022年3期
李龙沁 许光耀 高越 莫训强 李洪远





摘 要:归化是外来植物越过一系列障碍成为入侵植物的基本前提,對归化植物的种类组成及分布规律进行研究,有助于预防和管控植物入侵。该文基于文献报道和标本记录,对中国西部地区共12省(区)126个地级市的归化植物进行统计(包含所有维管束植物),并分析了这些植物的组成和时空分布特征。结果表明:(1)西部地区共有归化植物826种,分属107科473属,约占全国归化植物总数(1 099种)的75.16%。(2)时间上,根据回归拟合,15世纪前,西部地区的归化植物累积增长平稳,随后呈Logistic模式增长,1936年增长速率最大。目前,增长速度趋缓,但无意引入比例显著高于其他时期。(3)空间上,物种数及密度均呈自西北向东南、内陆向边境递增的趋势,且存在“强省会”现象。(4)回归树分析表明,年降水量、国内生产总值(GDP)、年均气温和耕地比例是西部地区归化植物分布格局的主要驱动因素。该研究结果丰富了西部地区归化植物的基础资料,有助于摸清西部地区外来植物入侵现状,并为其综合管控提供科学依据。
关键词: 生物入侵, 西部地区, 归化植物, 时空分布格局, 驱动因素
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2022)03-0429-11
Temporal and spatial distribution characteristics of
naturalized plants in West China
LI Longqin1,2, XU Guangyao1,3, GAO Yue1,2, MO Xunqiang4, LI Hongyuan1,2*
( 1. College of Environmental Science and Engineering, Nankai University, Tianjin 300350, China; 2. Key Laboratory of Environmental Technology
for Complex Trans-Media Pollution, Tianjin 300350, China; 3. Tianjin Huanke Environment Consul Ting Co., Ltd., Tianjin 300191,
China; 4. School of Geographic and Environmental Sciences, Tianjin Normal University, Tianjin 300387, China )
Abstract:Naturalization is the basic premise for exotic plants to surmount a series of obstacles and become invasive plants. It is helpful to prevent and control plant invasion for studying the species composition and distribution of naturalized plants. Based on literature reports and specimen records, we firstly maked statistics on naturalized plants in 126 prefecture-level cities in 12 provinces (regions) in West China, included all vascular plants, and analyzed the plant composition and temporal and spatial distribution characteristics. The results were as follows: (1) There were 826 species of naturalized plants belonging to 107 families and 473 genera in the western region, accounting for about 75.16% of 1 099 species of naturalized plants in the country. (2) In terms of time, according to regression fitting, the cumulative growth of naturalized plants in the western region was stable before the 15th century, and then showed a Logistic pattern of growth, and the growth rate was the largest in 1936. At present, the growth rate slowed down but the proportion of unintentional introduction was significantly higher than that in other periods. (3) Spatially, the number and density of species were increasing from northwest to southeast and inland to the border, and there was a phenomenon of strong provincial capitals. (4) Regression tree analysis showed that annual precipitation, GDP, average annual temperature and cropland ratio were the main driving factors for the distribution pattern of naturalized plants in the western region. These results enriches the basic data of naturalized plants in the western region, are conducive for finding out the current situation of alien plant invasion in the western region, and provide scientific basis for its comprehensive control.
Key words: biological invasion, western region, naturalized plants, temporal and spatial distribution pattern, driving factor
生物入侵已成為全球四大环境问题之一,对生态环境、人类健康及社会经济造成了严重威胁(王方浩等,2002;闫小玲等,2014;Hassan & Mohamed, 2020)。据不完全统计,我国每年因外来入侵物种而造成的经济损失超过2 000亿元(李大林,2014)。生物入侵还是导致全球生物多样性减少的最主要因素之一,入侵植物会通过占据优势地位降低本地植物的丰富度和多样性,甚至会沿着食物链或食物网影响本地动物的多样性,进而影响生态系统的运行过程(Eviner et al., 2012;许光耀等,2018)。20世纪80年代后,入侵物种危害日益猖獗,我国开始重视对外来入侵种的研究与防治。然而,由于国内区域生态调查不平衡,尤其是西部地区,其起始时间和调查程度差异较大,因此,西南地区较西北地区调查频率更高、起步早,记录更加详尽。云南是最早开始研究入侵植物问题的地区之一,如紫茎泽兰(Ageratina adenophora)的相关研究(薛纪如等,1979)。随后,广西(唐赛春等,2008)、贵州(韦美玉等,2006)、重庆(孙娟等,2009)等地也开展了对入侵植物的本底调查,而同一时期的西北地区,如新疆(张源,2007)等地区本底情况并不清楚,且对其危害也不够重视。目前,甘肃、宁夏和青海等部分地区仍缺少对入侵植物本底调查的基础数据,这使得区域入侵植物清单编制存在困难。
不同学者对外来植物的入侵状态属于逸生、归化或入侵的判断不同,目前仍然缺少认定的入侵标准,难以形成普遍认同的入侵植物名录(Van Kleunen et al., 2015),这使得明确并完善我国归化植物清单尤为重要。归化植物是指无需人类帮助(有可能存在人类干扰),能在多个生命周期内维持自我更新种群的外来植物,一般来说,能维持十年或广泛分布的植物就能经受各种环境因子筛选,即可被视为归化植物(Blackburn et al., 2011)。与入侵概念相比,归化植物的概念更加明确(Pyek et al., 2017)。Van Kleunen等( 2015)建立并编制了全球归化植物数据库(GloNAF数据库)及清单,共列入归化维管植物13 168种,占已知的所有维管植物的3.9%(Vinogradova et al., 2018),但其中中国地区的数据与实际情况存在较大误差。其后,越来越多的国家制定了归化植物清单(如印度、加纳、俄罗斯等)(Inderjit et al., 2018;Vinogradova et al., 2018;Ansong et al., 2019)。国内对于归化种研究起步较晚,闫小玲等(2019)和许光耀等(2019)分别在省级层面上对中国归化植物做了梳理,但数据的收集和更新均存在一定的差异性和滞后性。
越清晰的本底资料越有利于后续研究及安全管理,但当前绝大多数归化植物分布记录只精确到省级,极少统计到地市级。据此,本文在地级城市尺度上对西部地区12个省(市、自治区)所有地级市的归化植物进行统计,除嘉峪关市、金昌市、自贡市、南充市、乌海市5市数据缺失外,共获得126个地级市的归化植物的分布信息。对其种类组成结构及来源(包括原产地和引入用途),特别是时空分布格局进行了分析,以梳理西部地区归化植物的本底资料,揭示归化植物的分布规律及其驱动因素,为归化植物的深入研究和科学管理提供可靠的依据。
1 研究区域与研究方法
1.1 研究区域
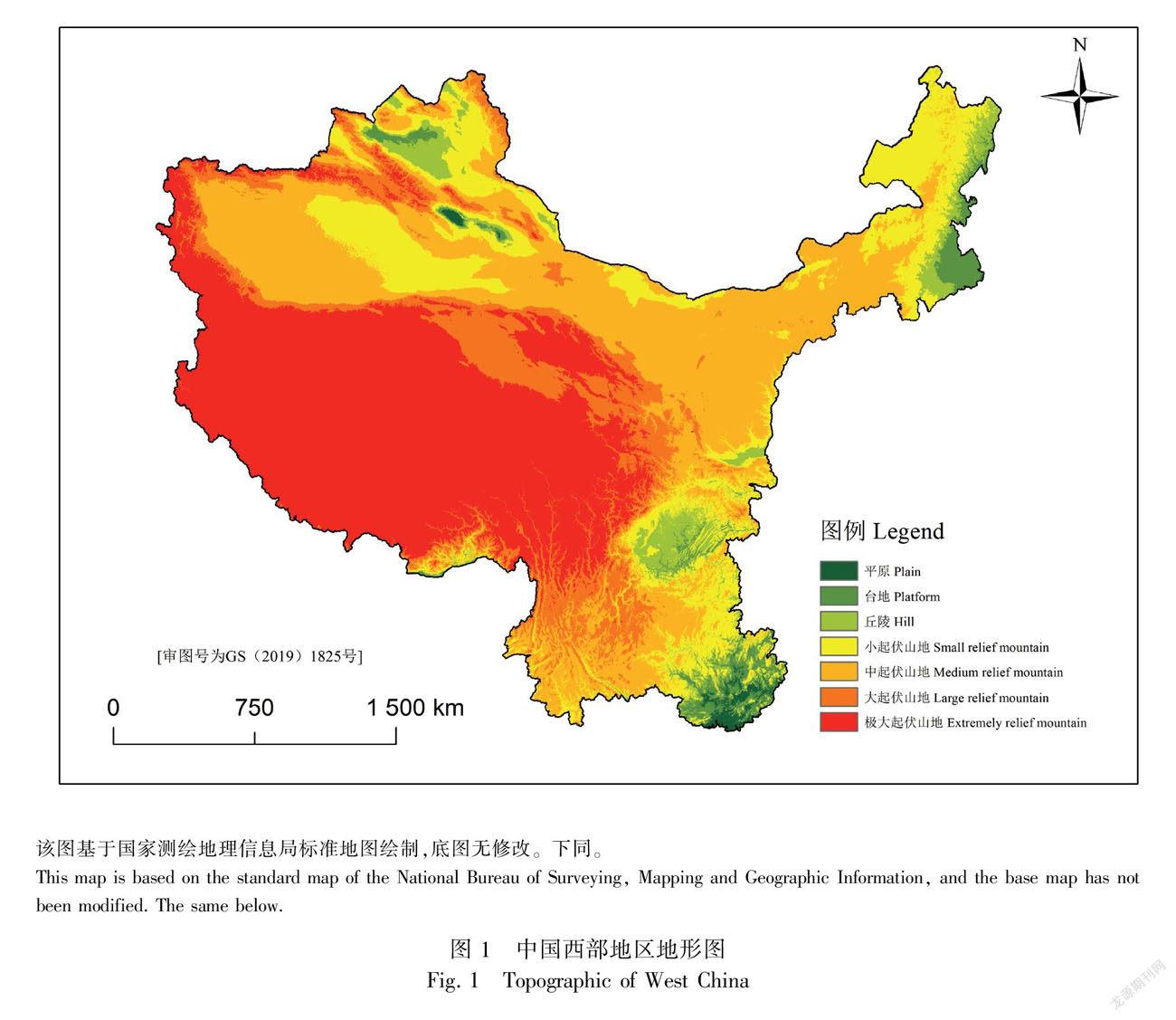
本研究区域是中国四大经济分区之一的西部地区(图1),包含陕西省、甘肃省、青海省、宁夏回族自治区、新疆维吾尔族自治区、西藏自治区、 四川省、重庆市、贵州省、云南省、内蒙古自治区和广西壮族自治区12个省(市、自治区),共126个市(盟、地区、自治州)。西部地区地域辽阔,地形特殊,自然景观丰富,人口稀少,经济欠发达,共与13个国家接壤,陆地边境线约1.8万km。气温和降水受青藏高原影响,均从西北向东南递增。土地面积约681.6万km2,占全国总面积的71.4%,人口数量约3.8亿,占全国总人口的27.2%。
1.2 研究方法
归化植物种类、分布数据及其他信息分别来源于《中国植物志》英文修订版(FOC,http://www.iplant.cn/)、The Plant List(TPL,http://www.theplantlist.com/)、自然标本馆(CFH,http://www.cfh.ac.cn/)、中国数字植物标本馆(CVH,http://www.cvh.ac.cn/)、中国植物图像库(PPBC,http://ppbc.iplant.cn/)、台湾物种名录(TaiCoL,http://taibnet.sinica.edu.tw/)、全球外来归化植物数据库(GloNAF,https://glonaf.org/)等网站,各省植物志及其他文献资料,共查询20 499个标本数据。通过逐一查阅、收集和核实,到形成最终的西部地区归化植物名单,并对植物中文名、拉丁名、科、属、原产地、引入时间、引入用途和分布地进行整理。其中,中文名(科、属、种)依据FOC确定,拉丁名以TPL确定,采用APG IV系统;原产地划分依据生物多样性信息标准(TDWG,https://www.tdwg.org/)的大洲分区体系;引入时间是指该种植物在中国的首次记录时间或首次引入时间;引入用途归纳为园林绿化、食用、材用、药用和无意引入5种。
自然环境数据包括年均气温、年降水量、干燥度指数、湿润度指数、0 ℃有效积温、归一化植被指数(NDVI)、草地比例、森林比例、水体比例、沙土比例和黏土比例,共11种,主要表示植物生长所需的不同环境及营养成分,分别从世界气象网(http://worldclimate.org/bioclim)、自然环境数据云平台(http://www.resdc.cn/Default.aspx)、国际农业研究磋商组织(www.cgiar-csi.org)、中国气象网(http://data.cma.cn)获得。社会环境数据主要反映区域经济发展整体水平、 交通运输业发展程度等人类活动干扰强度,包括GDP、人口密度、常住人口数、公路里程、公路货运量、夜间灯光指数、土地利用程度、土地利用动态度和聚落、耕地比例,共10种,均从中国社会与经济发展数据库(http://data.cnki.net)中获得。
1.3 数据分析
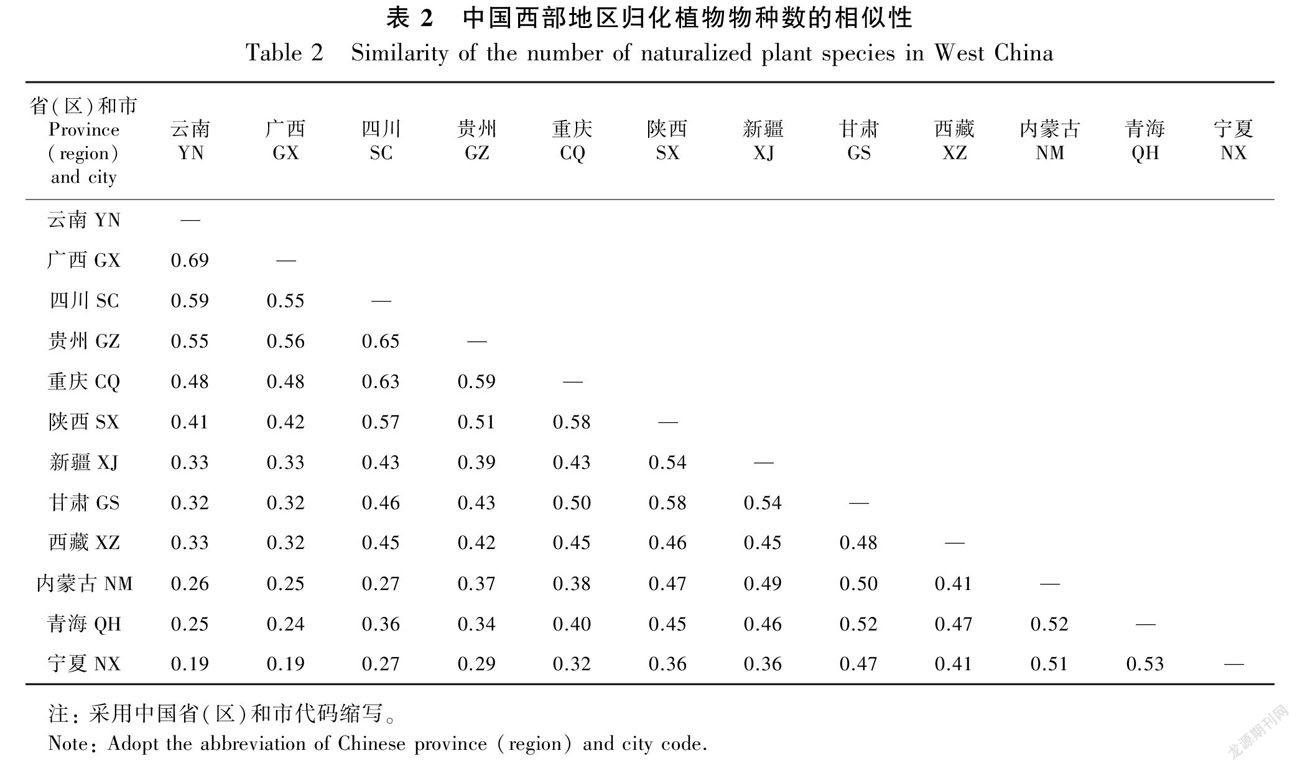
为探究归化植物随时间的累积分布情况,对西部地区归化植物累积数量与引入时间分别进行线性及Logistic拟合。为了解各科属在地级市的分布情况以及引入时间与原产地、引入用途的关系,通过Grubbs’s Test的方法对其显著性进行检验。此外,为分析其空间分布格局,在ArcGIS中采用自然间断点分级法(Jenks)对各省(区)、市物种数及密度进行分类,用Jaccard相似性指数反映各省(区)内的物种相似性。
J=ca+b-c (1)
式中:a和b表示两省(区)、市的物种数;c表示两省(区)、市之间的共有物种数。
根据各地级市的自然和社会环境数据,用Pearson系数法对126个地级市进行分析,去除相关性较强的因子,包括湿润度指数、有效积温、NDVI、沙土比例、黏土比例、常住人口数、公路货运量及土地利用程度8个因子,其余13个用于构建分类回归树。在分类回归树中,将面积作加权因子,根据CART算法在R中进行(De’Ath & Fabricius, 2000; Loh, 2011),以确定归化植物物种数与自然及社会因素之间的关系。
2 结果与分析
2.1 组成结构及来源
截止到2019年,共统计到中国共有归化植物112科578属1 099种(许光耀等,2019)。西部地区归化植物共统计到107科473属826种,约占全国归化植物总数的75.16%。从科的构成来看,菊科(Asteraceae)114种、豆科(Fabaceae)106种、禾本科(Poaceae)88种,三大优势科共计308种,占总种数的37.24%。此外,大于10种的科有13个(含269种),2~9种的科有53个(含275种),单种科共38个。从属的构成来看,物种数最多的是大戟属(Euphorbia)20种,大于10种的属共4个,6~9种的属共11个(含153种),2~5种的属有138个(含355种),单种属共319个(表1)。
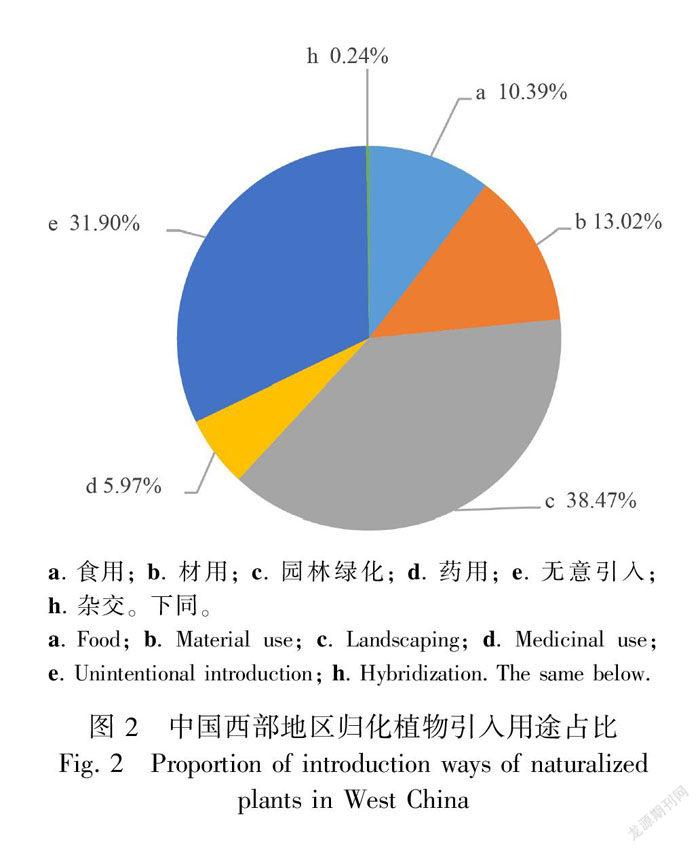
从引入用途看,园林绿化引入最多,共计322种(约占38.47%),无意引入共计267种,约占31.9%,食用、材用、药用等分别占总引入数的10.39%、13.02%、5.97%(图2)。
除10种植物原产地不详、11种杂交起源、6种栽培起源外,原产于美洲的归化植物最多,其中,北美约占21.42%,南美约占20.70%,此后依次为欧洲、温带亚洲、非洲、热带亚洲,大洋洲和太平洋岛屿起源较少。中国西南地区原产地分布趋势与西部地区及全国趋势一致(许光耀等,2019),原产于北美和南美最多,共占44.62%,而西北地区归化物种原产于温带亚洲和欧洲的物种最多,分别占22.37%和23.14%(图3)。
2.2 时间累积特征
共查询到700种归化植物的引入时间,约占总数的84.64%,其余126种绝大多数应在15世纪前引入。时间上,根据查询获得的700种引入时间统计发现,最早可追溯到公元前200年(先秦),变化规律分为两段(图4):第一段,从先秦至15世纪,西部地区归化植物累积速率较平缓,符合线性拟合(R2=0.985),共累积92种;第二段,在15世纪后,累积趋势符合Logistic曲线(R2=0.994),百余年间累积600余种。归化植物累积增长速率在清末迅速增长(EC20=1895),于民国时期(EC50=1936)达到最大,改革开放前期趋于稳定(EC80=1977)。
根据线性拟合和Logistic拟合分析出的四个时间节点,将引入时间分为五个时间段,对其原产地和引入用途进行分析(图5)。从原产地来看,15世纪前,原产温带亚洲的归化植物最多(约占31.37%),其次是欧洲(约占23.53%)。15世纪后,随着新大陆的发现导致原产于美洲的归化种数量快速增加。G检验发现,与其余时间段相比,1400年以前,原产于温带亚洲的比例显著高于其他地区(P<0.05),其余时间段各原产地比例相差不显著(P>0.05)。
从引入用途来看,1400年以前,因食用引入的物种最多(36.56%),如葡萄(Vitis vinifera)、茄(Solanum melongena)、西瓜(Citrullus lanatus)、芝麻(Sesamum indicum)等。1400—1977年,园林绿化引入的归化植物比例达52.22%,1977年后无意引入物种最多。G检验发现,1977年后,无意引入占比显著高于其他途径(P<0.05),其余各时间段引入用途的相差不显著(P>0.05)。
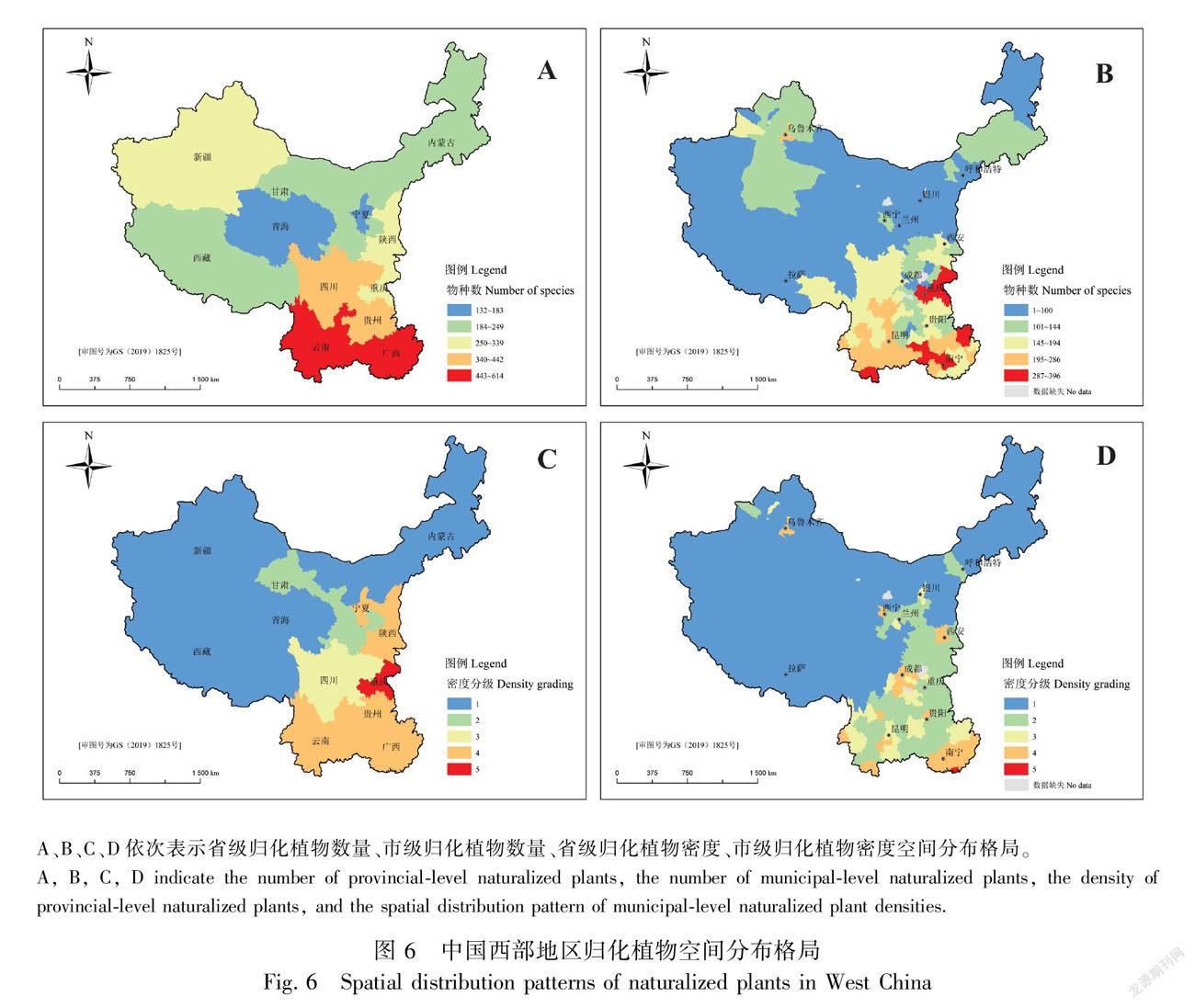
2.3 空间分布特征
空间上,西部地区归化植物物种数呈自内陆向边境、自西北向东南递增的趋势(图6:A,B;表2)。物种数最多的省份是云南,共614种,占西部地区归化植物总物种数的74.33%,其次是广西,共579种;最多的地级市为西双版纳傣族自治州,共396种,约占47.94%,其次是桂林市360种和南宁市342种。物种数最少的是宁夏和青海,分别为132种和183种;地级市为那曲地区、石嘴山市和阿里地区,分别为62种、70种和70種。此外,从省内的Jaccard相似性指数来看(表2),物种数最多与最少省份相似性指数为0.19,整体存在由物种数高的地区向物种数低的地区扩散的趋势。物种数最多的云南与广西,其相似性指数高达0.69,物种数最少的宁夏和青海的相似性指数也达0.53,说明存在相邻区域相似性高的趋势。
归化植物密度(物种数与面积之比)呈自西北向东南的递增趋势,(图6:C,D)。密度最高的是重庆(41.16种·万km-2);其次是广西(24.39种·万km-2)和贵州(22.20种·万km-2),地级市是北海市(578.4种·万km-2)和防城港市(306.2种·万km-2);内蒙古(1.71种·万km-2)和那曲地区(1.4种·万km-2)是密度最小的区和地级市。从省(区)内分布来看,陕西、甘肃、青海、宁夏、新疆、西藏、贵州、内蒙古8省(区)的省会城市密度高于省(区)内其他地级市。
2.4 空间分布影响因素分析
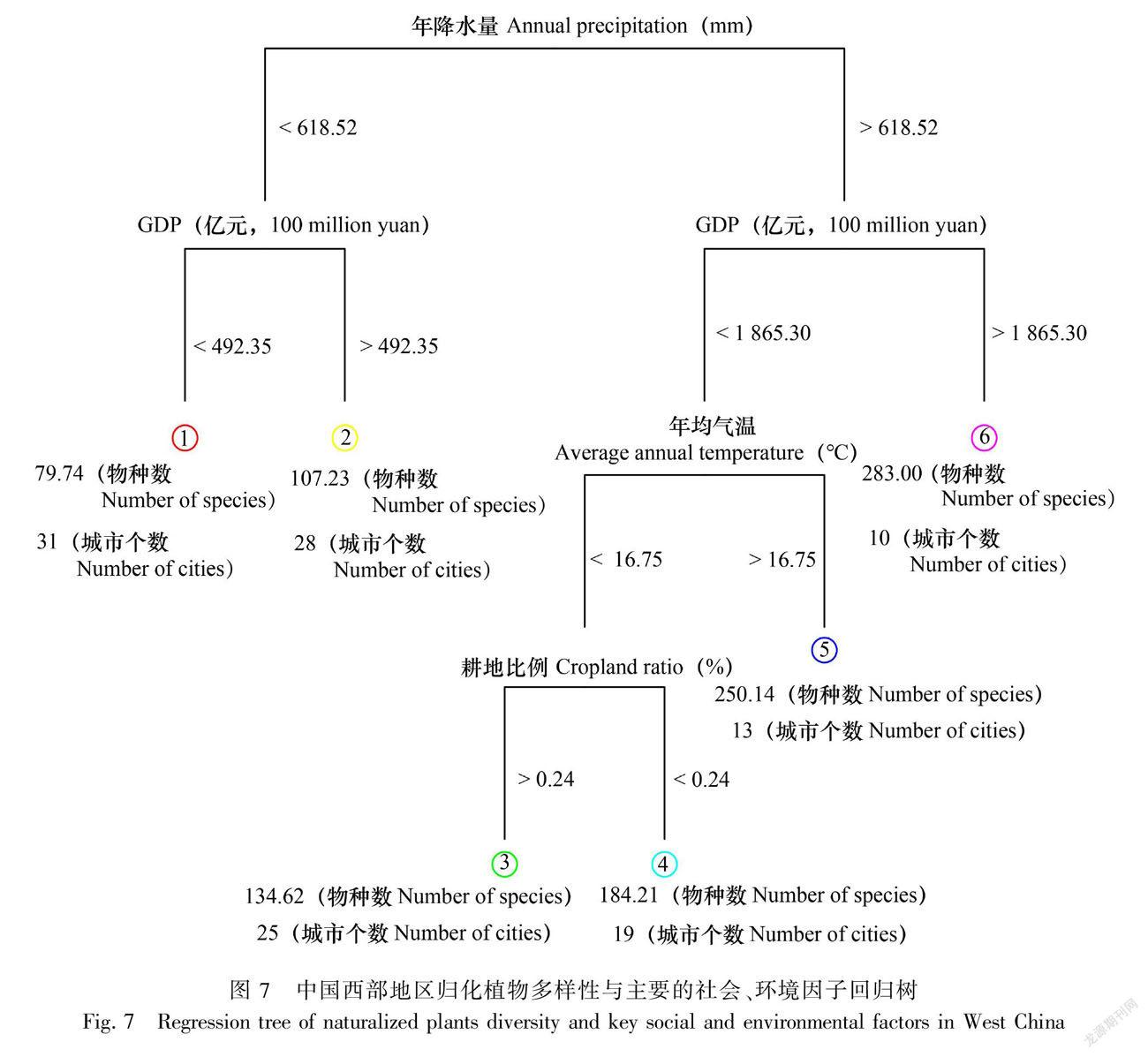
分类回归树分析表明,年降水量、GDP、年均气温和耕地比例是影响归化植物空间分布格局的主要因素。年降水量越小的地区归化物种数越低,GDP越高的地区归化物种数越高。当年降水量小于618.52 mm且GDP小于492.34亿元时,归化物种数最少(平均约79.74种),如海东地区、阿勒泰地区等;降水量较高且经济发达的地区归化物种数最多(平均约283.00种),如西安市、成都市、昆明市等;在降水量较高且经济欠发达地区,年均气温成为影响归化植物数量的关键因素,气温越高的地区归化物种数越高,当年均气温小于16.75 ℃时,平均归化物种数为250.14种,如西双版纳傣族自治州、梧州市、北海市等;年均气温小于16.75 ℃时,耕地比例越高的地区物种数低(图7)。
3 讨论与结论
3.1 时间累积特征分析
15世纪前,外来植物的传入途径较少, 人类活动干扰程度较低,西部地区归化植物累积较少,原产于温带亚洲的比例显著高于其他时期,但累积速率平稳增长。陆上丝绸之路增加了传入食用或无意引入的归化植物。据统计,中国现有的农作物中半数以上为外来植物,而在明朝(1368年)以前,大部分是通过陆上丝绸之路传入(刘启振和王思明,2016)。此外,其多数原产于亚洲西部地区及地中海沿岸,来自欧亚大陆的归化植物有着相似的生活史,能快速适应新的生长环境。
15世纪后,西部地区归化植物因园林绿化引入的比例近半数。随着通商口岸及殖民地的增加,由于交通运输、经济交流以及殖民者自身需求,引入了大量外来植物,使西部地区归化植物增长速率在清末开始爆发式增长(EC20=1895),并在民国时期(EC50=1936)达到最大。国内植物学研究的发展和中国植物学会的成立(肖蕾,2014),使大量植物信息被集中收集和记录,虽然会造成一定的统计偏差, 但也促使了其增长速率于1936年达到最大值。值得注意的是,1977年后,随着改革开放,西部地区的经济逐渐发展,人口流动增大,其无意引入占比也显著高于其他途径。然而,较早归化的植物会占据大量空余生态位,阻止了其他植物归化,虽使得1977年后增长速率逐步趋于稳定(EC80=1977),但仍然有归化的风险。这与前人研究结果一致,一定时期归化植物增长速率与当时经济发展状况有密切的联系(Cabra-Rivas et al., 2016),贸易活动是影响外来植物入侵和扩散的重要因素(蒙彦良和陈凤新,2020)。
进入21世纪,根据Logistic模型显示,当前西部地区归化物种处于增长速率逐步稳定的阶段。国家检验检疫政策逐步健全,对动植物检疫更加严格,植物引种逐渐规范,使有意引入和驯化的归化植物增长速率降低(何佳瑶等,2019)。虽然植物检疫工作在近年来取得一定成效,对抵御外来生物入侵起着重要作用,但仍然存在的入侵风险不容忽视。此外,随着全球化的进一步发展,“一带一路”政策的实施以及国际贸易往来的进一步加强,西部地区的经济发展必将迎来新的局面 (任保平等,2019)。经济及交通运输网络的发展,人口流动及对外交流的增加等愈发频繁的人类活动,也会增加有意或无意引入外来种,从而可引发归化植物累积速度再次加快的风险,为植物入侵提供可乘之机。
3.2 空间分布特征分析
从12个省(区)分布看,有8个省会城市归化植物密度最高,表现出明显的强省会现象。省会城市中,园林绿化是归化植物最主要的引入用途,同一般城市相比,省会城市具有更完善的基础设施和更丰富的资源,城市景观建设中园林植物的应用更加丰富,受人类活动影响更大,在获得更多资源的同时也给植物归化带来更多的机会。
港口、口岸及边界城市拥有特殊的地理位置,丰富的旅游资源和频繁的国际交流等条件,增加了植物归化的可能性(Foxcroft et al., 2017),这也是北海市和防城港市归化种密度最高的原因。值得注意的是,西双版纳傣族自治州位于全球生物多样性热点地区之一的Indo-Burma内(Myers et al., 2000),作为中国陆地生物多样性保护的关键地区(Tang et al, 2006),也是西部地区归化植物数量最多的地级市,占西部地区总数近50%。其原因除了独特的地理位置和适宜的气候条件外,其中的绝大多数外来植物是由于植物园的长期引种而归化,位于勐腊县境内的中国科学院西双版纳热带植物园是西双版纳引种外来植物最多的机构(李园等,2006)。植物园所在地会成为归化植物的分布热点区域,国内植物园的平均物种数在3 000左右,西双版纳热带植物园约5 000种,其中有1/3是外来植物(He, 2002)。植物的收集、引种和驯化是植物园的重要任务,长期引种使外来植物丰富了植物物种多样性,但同时也带来了对生态环境的危害。因此,各省会、港口城市需重点防范,植物园引种应更加警惕。
与前人研究结果一致,气候和人类干扰对生物入侵进程的影响尤为明显(吴昊和丁建清,2014)。回归树分析表明,年降水量、GDP和年均气温是主要驱动因素。耕地比例的增高會阻碍外来植物的归化,一般来说耕地多的地方欠发达,干扰相对较弱,且几千年的耕种过程中,杂草类植物早就占据了作物之外的空余生态位,阻止了其他植物的归化(Van Kleunen et al., 2015)。
总体来说,我国西部地区地形特殊,青藏高原的隆起形成了独特的高原气候,直接影响了降水量,形成西北的干旱气候(范广州和程国栋,2003),海拔高的地区人口稀少,人类活动影响较小,使归化植物数量较少。某种程度上来说,地形是影响西部地区归化植物数量最重要的因素,值得在后续研究中对其进一步深化。宁夏、青海、新疆等地区调查频率较少,调查区域往往受时间、频率、交通等影响(Hyndman et al., 2015),信息收集不够全面,植物标本也存在物种鉴定错误、信息记录不详等问题,导致本文统计结果存在一定的不确定性,需在今后的研究中持续完善。
综上所述,中国西部地区共统计到归化植物826种,原产于美洲的最多,园林绿化是主要的引入用途。西部地区归化植物空间分布整体上呈西北向东南、内陆向边境递增的趋势,年降水量和GDP是其最主要的影响因素,景观建设、对外贸易、大型植物园和调查频次会影响归化植物省内空间分布格局。当前西部地区的归化物种累积速率趋于平缓,但随着“一带一路”政策的实施和经济的发展,西部地区尤其是省会城市归化植物累积速度可能会增快,相关部门仍需保持警惕。此外,西部地区特殊的地形, 在一定程度上直接影响降水、 气温及人口分布,其对归化植物累积的影响值得进一步探讨。调查较少而数据缺乏的地区(如宁夏、青海等地)需增加调查频次,以降低统计误差,为西部地区归化物种名录编制及植物入侵的预测、预防及管理提供更准确的依据。
参考文献:
ANSONG M, PERGL J, ESSL F, et al., 2019. Naturalized and invasive alien flora of Ghana[J]. Biol Invasions, 21(3): 669-683.
BLACKBURN TM, PYEK P, BACHER S, et al., 2011. A proposed unified framework for biological invasions[J]. Trend Ecol Evol, 26(7): 333-339.
CABRA-RIVAS I, SALDANA A, CASTRO-DIEZ P, et al., 2016. A multi-scale approach to identify invasion drivers and invaders’ future dynamics[J]. Biol Invasions, 18(2): 411-426.
DE′ATH G, FABRICIUS KE, 2000. Classification and regression trees: a powerful yet simple technique for ecological data analysis[J]. Ecology, 81(11): 3178-3192.
EVINER VT, GARBACH K, BATY JH, et al., 2012. Measuring the effects of invasive plants on ecosystem services: challenges and prospects[J]. Invasive Plant Sci Manag, 5(1): 125-136.
FAN GZ, CHENG GD, 2003. Reason analysis of the influence of Qinghai-Xizang Plateau uplifting on arid climate forming in northwest China (I): Influence on general circulation of atmosphere[J]. Plateau Met, 22(S1): 45-57. [范廣洲, 程国栋, 2003. 青藏高原隆升对西北干旱气候形成影响的模拟(I):对大气环流影响[J]. 高原气象, 22(S1): 45-57.]
FOXCROFT LC, PYEK P, RICHARDSON DM, et al., 2017. Erratum to: plant invasion science in protected areas: progress and priorities[J]. Biol Invasions, 19(8): 2503-2505.
HASSAN MO, MOHAMED HY, 2020. Allelopathic interference of the exotic naturalized Paspalum dilatatum Poir. threatens diversity of native plants in urban gardens[J]. Flora, 266(2020): 151593.
HE JY, CHEN K, PAN XB, et al., 2019. Analysis of plant pest interception at China’s ports of entry during 2016-2017[J]. Plant Quarant, 33(6): 34-37. [何佳遥, 陈克, 潘绪斌, 等, 2019. 2016—2017年中国进境植物疫情截获情况分析[J]. 植物检疫, 33(6): 34-37.]
HE SA, 2002. Fifty years of botanical gardens in China[J]. Acta Bot Sin, 9(44): 1123-1133.
HYNDMAN RJ, MESGARAN MB, COUSENS RD, 2015. Statistical issues with using herbarium data for the estimation of invasion lag-phases[J]. Biol Invasions, 17(12): 3371-3381.
INDERJIT, PERGL J, VAN KLEUNEN M, et al., 2018. Naturalized alien flora of the Indian states: biogeographic patterns, taxonomic structure and drivers of species richness[J]. Biol Invasions, 20(6): 1625-1638.
LI DL, 2014. China’s annual economic losses due to alien biological invasion exceed 200 billion yuan[J]. Guangxi Qual Super Guide Periodical, (11): 30. [李大林, 2014. 我国每年因外来生物入侵经济损失超两千亿元[J]. 广西质量监督导报, (11): 30.]
LI Y, WU ZL, LI LS, et al., 2006. A preliminary study on species diversity usage and ecological impact of alien plants in Xishuangbanna[J]. J Plant Resour Environ, 15(2): 68-72. [李园, 吴兆录, 李丽莎, 等, 2006. 西双版纳外来植物的物种多样性、用途和生态危害的初步研究[J]. 植物资源与环境学报, 15(2): 68-72.]
LIU QZ, WANG SM, 2016. The extraterritorial crops introduced into China via the Silk Road[J]. Chin Wild Plant Resour, 35(6): 5-20. [劉启振, 王思明, 2016. 陆上丝绸之路传入中国的域外农作物[J]. 中国野生植物资源, 35(6): 5-20.]
LOH WY, 2011. Classification and regression trees[J]. WIREs: Data Mining Knowl Discov, 1(1): 14-23.
MENG YL, CHEN FX, 2020. Changes and growth forecast of invasive plants in China from the perspective of trade[J]. Plant Quarant, 34(2): 1-8. [蒙彦良, 陈凤新, 2020. 贸易视角下中国外来植物的变化及其增长预测[J]. 植物检疫, 34(2): 1-8.]
MYERS N, MITTERMEIER RA, MITTERMEIER CG, et al., 2000. Biodiversity hotspots for conservation priorities[J]. Nature, 403(6772): 853-858.
PYEK P, PERGL J, ESSL F, et al., 2017. Naturalized alien flora of the world: Species diversity, taxonomic and phylogenetic patterns, geographic distribution and global hotspots of plant invasion[J]. Preslia, 89(3): 203-274.
REN BP, ZHANG Q, 2019. Achievements, experiences and transition of economic development in the western region in 20 years of development of western China[J]. J Shaanxi Norm Univ(Philos Soc Sci Ed), 48(4): 46-62. [任保平, 张倩, 2019. 西部大开发20年西部地区经济发展的成就、经验与转型[J]. 陕西师范大学学报(哲学社会科学版), 48(4): 46-62.]
SUN J, YANG GF, CHEN YC, et al., 2009. Exotic plants in the Jinfo Mountain Nature Reserve: Species ascertain and their distribution[J]. Acta Pratacul Sin, 18(3): 34-42. [孙娟, 杨国峰, 陈玉成, 等, 2009. 金佛山自然保护区外来入侵植物种及其分布情况[J]. 草业学报, 18(3): 34-42.]
TANG SC, L SH, HE CX, et al., 2008. The alien invasive plants in Guangxi[J]. Guihaia, 28(6): 775-779. [唐赛春, 吕仕洪, 何成新, 等, 2008. 广西的外来入侵植物[J]. 广西植物, 28(6): 775-779.]
TANG ZY, WANG ZH, ZHENG CY, et al., 2006. Biodiversity in China’s mountains[J]. Front Ecol Environ, 4(7): 347-352.
VAN KLEUNEN M, DAWSON W, ESSL F, et al., 2015. Global exchange and accumulation of non-native plants[J]. Nature, 525(7567): 100-103.
VINOGRADOVA Y, PERGL J, ESSL F, et al., 2018. Invasive alien plants of Russia: insights from regional inventories[J]. Biol Invasions, 20(8): 1931-1943.
WAN FH, GUO JY, WANG DH, 2002. Alien invasive species in China: Their damages and management strategies[J]. Biodivers Sci, 10(1): 119-125. [萬方浩, 郭建英, 王德辉, 2002. 中国外来入侵生物的危害与管理对策[J]. 生物多样性, 10(1):119-125.]
WEI MY, LIU LP, WEN ZR, 2006. Growth and control of exotic plant in Qiannan areas of Guizhou[J]. Guizhou Agric Sci, 34(2): 35-38. [韦美玉, 刘丽萍, 文治瑞, 2006. 贵州黔南地区外来植物逸生机危害调查[J]. 贵州农业科学, 34(2): 35-38.]
WU H, DING JQ, 2014. Recent progress in invasion ecology[J]. Chin Sci Bull, 59(6): 438-448. [吴昊, 丁建清, 2014. 入侵生态学最新研究动态[J]. 科学通报, 59(6): 438-448.]
XIAO L, 2014. Botanical society of China during the period of the republic of China[J]. J Hebei N Univ (Soc Sci Ed), 35(3): 43-63. [肖蕾, 2014. 民国时期的中国植物学会[J]. 河北北方学院学报(社会科学版), 35(3): 43-63.]
XU GY, LI HY, MO XQ, et al., 2018. Research review on the advances of the invasive plants and the ecological effects of the related factors concerned[J]. J Safe Environ, 18(1): 375-380. [许光耀, 李洪远, 莫训强, 等, 2018. 入侵植物生态效应及其影响因素研究进展[J]. 安全与环境学报, 18(1): 375-380.]
XU GY, LI HY, MO XQ, et al., 2019. Composition and spatial-temporal distribution of Chinese naturalized plants[J]. Acta Phytoecol Sin, 43(7): 601-610. [许光耀, 李洪远, 莫训强, 等, 2019. 中国归化植物组成特征及其时空分布格局分析[J]. 植物生态学报, 43(7): 601-610.]
XUE JR, DONG SR, YIN JH, et al., 1979. Eupatorium coelestium L.: It’s adaptability, damage and control[J]. Acta Bot Yunnan, 1(1): 106-113. [薛纪如, 董世仁, 印嘉祜, 等, 1979. 紫茎泽兰的适应性能、危害情况及防除措施[J]. 云南植物研究, 1(1): 106-113.]
YAN XL, LIU QR, SHOU HY, et al., 2014. The categorization and analysis on the geographic distribution patterns of Chinese alien invasive plants[J]. Biodivers Sci,22(5): 667-676. [闫小玲, 刘全儒, 寿海洋, 等, 2014. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性, 22(5): 667-676.]
YAN XL, WANG ZH, MA JS, 2019. The checklist of the naturalized plants in China[M]. Shanghai: Shanghai Science and Technology Press: 1-425. [闫小玲, 王樟华, 马金双, 2019. 中国归化植物名录[M]. 上海: 上海科学技术出版社: 1-425.]
ZHANG Y, 2007.The exotic plants of Urumqi City[D]. Urumqi: Xinjiang Normal University: 1-67. [张源, 2007. 乌鲁木齐市外来植物研究[D]. 乌鲁木齐: 新疆师范大学: 1-67.]
(责任编辑 周翠鸣)
收稿日期:2021-01-29
基金项目:天津市重大科技专项与工程(18ZXSZSF00200) [Supported by Major Key Project and Engineering of Science and Technology and Projects in Tianjin(18ZXSZSF00200)]。
第一作者: 李龙沁(1997-),硕士研究生,研究方向为植物多样性和生态恢复,(E-mail)longq_lee@163.com。
通信作者:李洪远,博士,教授,研究方向为生态恢复、植被生态与湿地生态,(E-mail)eialee@nankai.edu.cn。
猜你喜欢
社会科学研究(2016年6期)2017-02-16
安徽农学通报(2017年1期)2017-02-15
现代营销·学苑版(2016年12期)2017-01-23
商业经济(2016年3期)2016-12-23
亚太教育(2016年35期)2016-12-21
人民论坛(2016年32期)2016-12-14
经济研究导刊(2016年28期)2016-12-14
经济研究导刊(2016年24期)2016-12-12
商场现代化(2016年21期)2016-10-21
科技视界(2016年15期)2016-06-30
