
南亚热带四种不同演化程度禾本科植物夏季光能利用策略差异分析
2022-04-08 01:08张雅芳王海玲朱师丹张小燕朱俊杰
广西植物 2022年3期
关键词:高温
张雅芳 王海玲 朱师丹 张小燕 朱俊杰


摘 要:为了探究甘蔗、地毯草、芦苇、佛肚竹这四种禾本科植物的光能利用策略,该文以大田或原生境植株为材料,于炎夏伏天最热时节活体监测其叶绿素荧光和光合气体交换特性,并分析其色素含量。结果表明:(1)C4甘蔗和C4地毯草具有高凈光合速率(Pn),是消耗利用光能的主要手段,并且高Pn和高水分利用效率(WUE)、高量子效率(Φi)耦合在一起;(2)C3芦苇也具有较高的Pn、WUE和Φi,其较高的类胡萝卜素与非光化学淬灭热耗散(NPQ)相结合消耗多余光能;(3)C3佛肚竹Pn、WUE和Φi值均最低,但具有最大的叶黄素库容和较高的叶黄素循环转换效率(DPS),通过叶黄素循环和NPQ的偶联消耗多余光能。此外,四种植物光能利用策略也与其生活型有密切关联。该研究结果对甘蔗、地毯草和佛肚竹培育具有重要的现实参考价值,对芦苇生境生态修复和生态规划有科学指导价值。
关键词: 光合演化, 光适应, 禾本科植物, 高温, 南亚热带
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2022)03-0440-10
Analysis on the differences of light utilization strategies of
four Poaceae species with different evolution degrees in the
south subtropical region of China during summer period
ZHANG Yafang1,2, WANG Hailing1,2, ZHU Shidan2, ZHANG Xiaoyan1,2, ZHU Junjie1,2*
( 1. Forestry College of Guangxi University, Nanning 530004, China; 2. State Key Laboratory of Protection and
Utilization Subtropical Agricultural Biological Resources, Guangxi University, Nanning 530004, China )
Abstract:In order to explore the light energy utilization strategy of four Poaceae species, including sugarcane, carpet grass, bulrush and buddha bamboo, the plants in the field or original habitat were used as materials. We determined the chlorophyll fluorescence and photosynthetic gas exchange characteristics from fresh seedlings leaves in vivo, and analyzed the pigment contents during the hottest period of the summer. The results were as follows: (1) The C4 sugarcane and carpet grass had high net photosynthetic rate (Pn) coupled with high water use efficiency (WUE) and high quantum yield efficiency (Φi), thus being able to assimilate a large fraction of light energy; (2) The C3 bulrush had relatively high Pn, WUE and Φi, and its high carotenoid pool which coupled high nonphotochemical quenching heat dissipation (NPQ) might help the bulrush to get rid of the excessive light energy; (3) The C3 buddha bamboo had the lowest Pn, WUE and Φi, however, its large carotenoid contents and high xanthophyll de-epoxidation level (DPS), coupled with high NPQ were expected to help the leaves to dissipate the excess light energy. In addition, the light-use strategies of the four species were also affected by their life forms. These findings are expected to help breeding programs for sugarcane, carpet grass and buddha bamboo, and to provide a physiological reference for wetland ecological restoration and planning of bulrush.
Key words: evolution of photosynthesis, photosynthetic adaptation, Poaceae species, hot stress, south subtropical
禾本科是被子植物“五大家族”之一,从炎热的荒漠到严寒的南极均有分布,覆盖了全球1/5以上的陆地面积,贡献了约1/4的陆地初级净生产力(Lloyd & Farquhar, 1994),生态和经济价值显著。
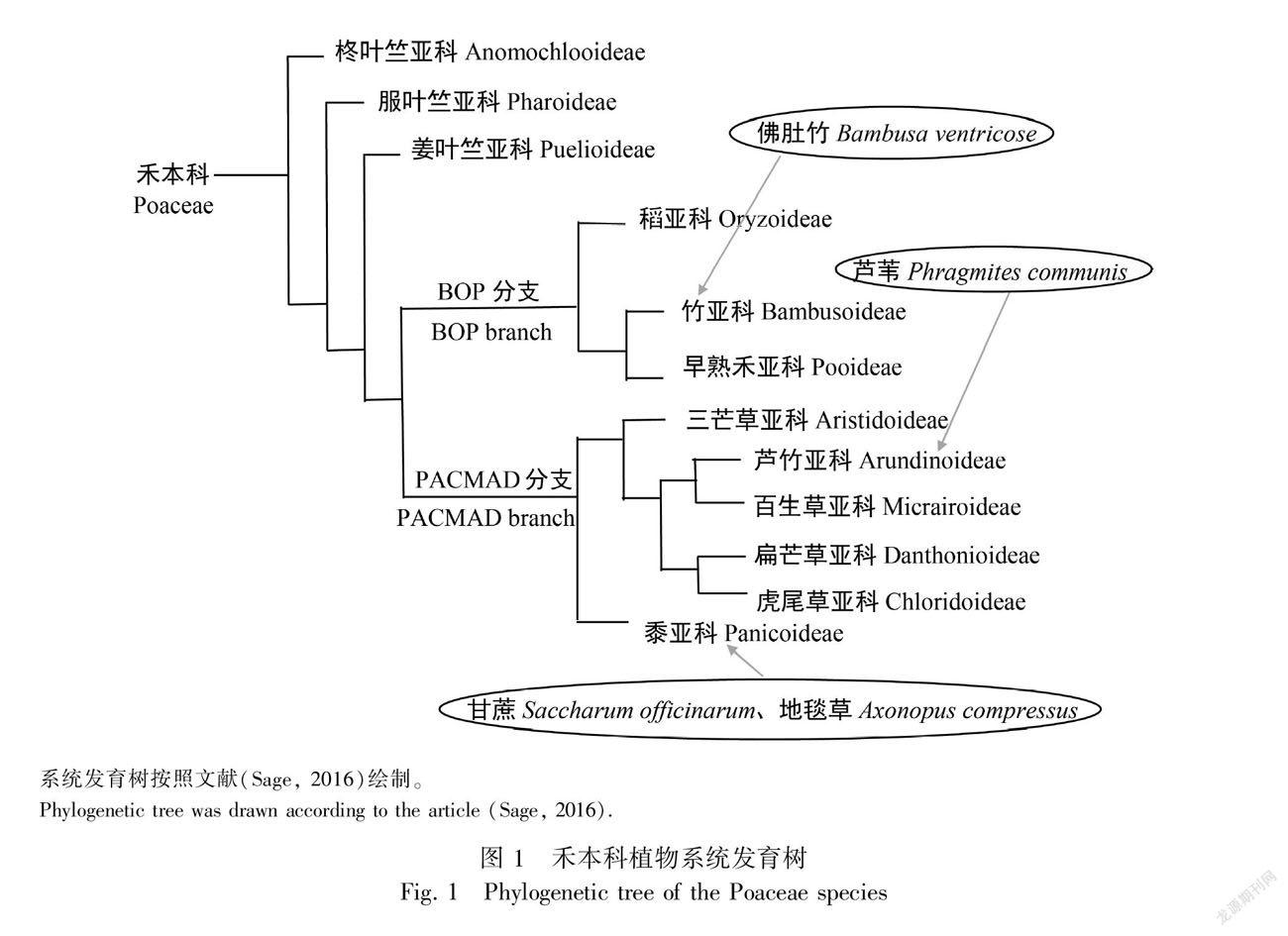
基于禾草系统发育工作组(Grass Phylogeny Working Group, GPWG)(GPWG I et al., 2001; GPWG Ⅱ et al., 2012)和后续学者(Cotton et al., 2015; Soreng et al., 2015)的修正系统,柊叶竺亚科、服叶竺亚科和姜叶竺亚科依次形成禾本科系统发育树的基系,在此基础上演化出BOP分枝(稻亚科、竹亚科、早熟禾亚科)和PACMAD分支(黍亚科、三芒草亚科、芦竹亚科、百生草亚科、扁芒草亚科、虎尾草亚科)。3 000万年前渐新世时期,低CO2浓度和干旱环境压力驱动部分禾本科植物从C3途径进化出C4途径,这是禾本科植物系统演化中的重要里程碑。目前,柊叶竺亚科、服叶竺亚科和姜叶竺亚科基系和BOP分支植物中尚未发现C4植物,PACMAD分支中的黍亚科、三芒草亚科、芦竹亚科、虎尾草亚科共有5 044种C4植物,约占禾本科总数的41%(Sage, 2016)。C4植物较C3植物有更高的光合速率和更强的生长能力,然而,随CO2浓度升高,全球气候变暖,C4植物的优势可能弱化而变为劣势(Edwards et al., 2010)。
光合机构吸收的光能超过光合作用所能利用的量时会引发光抑制,严重的光抑制导致光氧化、光漂白等损伤和破坏(Acebron et al., 2020)。植物在长期进化中形成一系列光抑制防御机制,例如:通过提高光合能力降低光能过剩(Savitch et al., 2002);依靠跨内囊体膜质子梯度的高能态淬灭耗能(Holt et al., 2005),其中PsbS蛋白(Li et al., 2000)和叶黄素循环(Horton et al., 1996)起了核心作用; 天线系统(Niyogi et al., 1998)和反应中心 (Matsubara & Chow, 2004) 淬灭耗能;如光呼吸(Ort & Barker, 2002)、水-水循环(Asada, 1999)和叶绿体呼吸(Nixon & Rich, 2007)等代谢耗能。目前,植物光抑制防御在模式植物和C3植物方面研究较多,并且这些研究主要聚焦于揭示光能利用的生理和分子机制,从生态视角比较不同类群,如C3和C4植物,湿生和旱生,高位芽挺立和地面匍匐植物光能利用策略差异的报道还很少。
我國南亚热带地区气候的一个显著特点是夏季漫长而湿热,然而该地区植物却在此时期生长最旺盛。它们适应这种气候的光合生理生态机制目前尚不清楚。为此,本研究以我国南亚热带地区重要代表性禾本科植物PACMAD分支中黍亚科高粱族甘蔗亚族甘蔗属的旱生C4甘蔗(Saccharum officinarum, SO)、黍亚科雀稗族雀稗亚族地毯草属的地生C4地毯草(Axonopus compressus, AC)、芦竹亚科蓝沼草族芦苇属的挺水C3芦苇(Phragmites communis, PC)和BOP分支中竹亚科簕竹族簕竹亚族簕竹属丛生型C3佛肚竹(Bambusa ventricose, BV)为材料(图1),研究它们在炎夏伏天的光能利用策略及差异,为植物光适应和禾本科植物光合演化相关研究提供新资料,同时为甘蔗、地毯草和佛肚竹生产提供参考,为芦苇生态修复和生态规划提供科学指引。
1 材料与方法
1.1 试验地点及材料
试验地点位于广西壮族自治区南宁市西郊广西大学(108°17′ E、22°50′ N)校园内。该地属我国南亚热带气候区,为湿润的亚热带季风气候,年均降水量1 304.2 mm,集中于5—9月。春暖夏热, 秋冬短暂,几乎全年无霜,年均温21.3 ℃, l月均温12.9 ℃, 7月均温28.3 ℃,土壤为砖红壤,pH值4.5~6.5。
甘蔗、地毯草、芦苇和佛肚竹四种试验材料均生长于广西大学东校园, 其中:甘蔗为旱地大田种植,芦苇为桶栽培养,低水肥常规管理,测定时株高约2 m;地毯草为校园绿化草坪植物,由工人定期维护,茎长10~15 cm;佛肚竹自然生长于水塘边,株高约为2 m。
1.2 叶绿素荧光参数的测定
在2020年7月10—17日连续晴朗炎热天气,选取生长健康的芦苇、甘蔗、佛肚竹最高可见肥厚带叶,地毯草直立枝顶端倒数第2叶,使用Dual-PAM-100叶绿素荧光仪(WALZ,Germany)活体测定所有叶绿素荧光参数。每天凌晨5:00测定经一整晚暗适应样叶的光系统Ⅱ (PS Ⅱ) 最大光化学效率(Fv/Fm)和光系统Ⅰ(PS Ⅰ)P700最大氧化量子产额(Pm)。上午8:00经暗适应30 min后开启1 178 μmol·m-2·s-1光化光进行诱导,达稳态后记录PS Ⅰ实际光合量子产额Y(I)、供体侧非光化学能量耗散额Y(ND) 、受体侧非光化学能量耗散额Y(NA)、PS Ⅱ实际光合量子产额Y(Ⅱ)、热耗散的量子产额Y (NPQ) 、组成型量子产额Y (NO)、光化学淬灭系数(qP)、热耗散系数(NPQ)以及PS Ⅰ和PS Ⅱ相对电子传递速率ETR I、ETR Ⅱ等参数。
1.3 光合气体交换参数的测定
于2020年7月10—17日的上午8:00—11:00(气温32~36 ℃,光强480 ~2 060 μmol·m-2·s-1),使用LI-6800便携式光合仪(Li-Cor, USA)测定用于叶绿素荧光参数测定的叶片光合气体交换参数。流速设为500 μmol·m-2·s-1,通过小钢瓶控制CO2浓度到400 μmol·mol-1,相对湿度60%,叶室温度32 ℃。测定时先在1 500 μmol·m-2·s-1光强下(红蓝光比例9∶1)诱导,达稳态后记录净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等参数,再按2 000、1 800、1 500、1 000、800、600、400、250、200、150、100、75、50、25、0 μmol·m-2·s-1的光梯度测光响应曲线,每个梯度下至少保持3 min。光响应曲线参数使用直角双曲线修正模型(Ye & Yu, 2008)进行拟合。内禀水分利用效率(WUEi)按公式WUEi = Pn/Gs计算,叶片瞬时水分利用效率(WUE)按公式WUE=Pn/Tr计算。
1.4 叶片色素含量及叶黄素循环组分测定
完成叶绿素荧光和光合气体交换参数的测定后,分别于凌晨和正午剪取测定叶片,参照陈敏氡等(2016)的方法,准确称取0.1 g鲜叶, 加液氮研磨,再加1 mL丙酮冰箱内避光浸提12 h,浸提液经5 000 r·min-1 3 ℃离心10 min,取上清液,添加1 mL丙酮避光低温浸提2 h,再次3 ℃离心10 min,最后将两次上清液合并,经0.22 nm有机膜过滤后上高效液相色谱仪(WATERS,ALLIANCE E2695,USA)测定色素含量。色谱条件参照朱俊杰(2007)的方法,色谱柱Waters Symmetry RP18(5 μm,4.6 mm,250 mm)购自北京鼎新昌盛科技有限公司,流动相A液为乙腈∶甲醇∶水(84∶9∶7),流动相B液为甲醇,流速为1 mL·min-1,采用梯度洗脱法,先用100% A液洗脱8 min,8~10 min后降为0% A液,10~20 min 0% A液,20~21 min上升为100% A液,21~38 min 100% A液,由Waters2998 PAD(WATERS,USA)检测器检测,检测波长445 nm。叶绿素b(Chlb)、叶绿素a(Chla)标准样购自Sigma 公司,新黄质(N)、叶黄质(L)、紫黄质(V)、环氧玉米黄质(A)、玉米黄质(Z)购自BOC Sciences公司。叶黄素循环转换率(DPS)按DPS =(A+Z)/(V+A+Z)计算。
1.5 数据分析与处理
各测定指标重复三次(n=3),结果以平均值±标准差表示,Duncan法比较四种植物的光合气体交换、叶绿素荧光参数及色素含量的差异(P<0.05),使用Microsoft Excel 2013软件处理数据,用SigmaPlot 14.0软件作图,利用SPSS 25.0软件统计分析。
2 结果与分析
2.1 四种禾本科植物两个光系统凌晨最大光化学效率
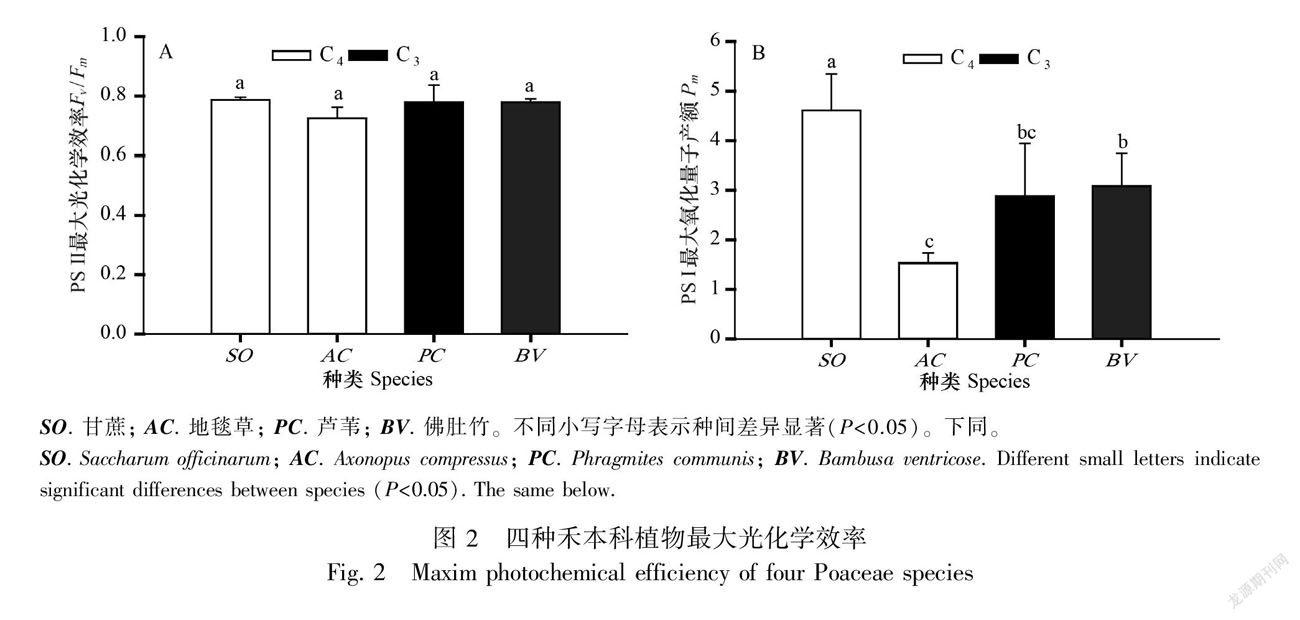
如图2:A所示,四种禾本科植物炎夏最热时段PS Ⅱ凌晨Fv/Fm值均在0.8以下,虽然统计结果显示种间没有显著差异(P>0.05),但是C4地毯草(0.727)表现出明显的光抑制。进一步分析PS Ⅰ的Pm值,种间差异(P<0.05)则突显出来,甘蔗最高,地毯草最低, C3的芦苇和佛肚竹处于中间(图2:B)。
2.2 四种禾本科植物两个光系统能量利用
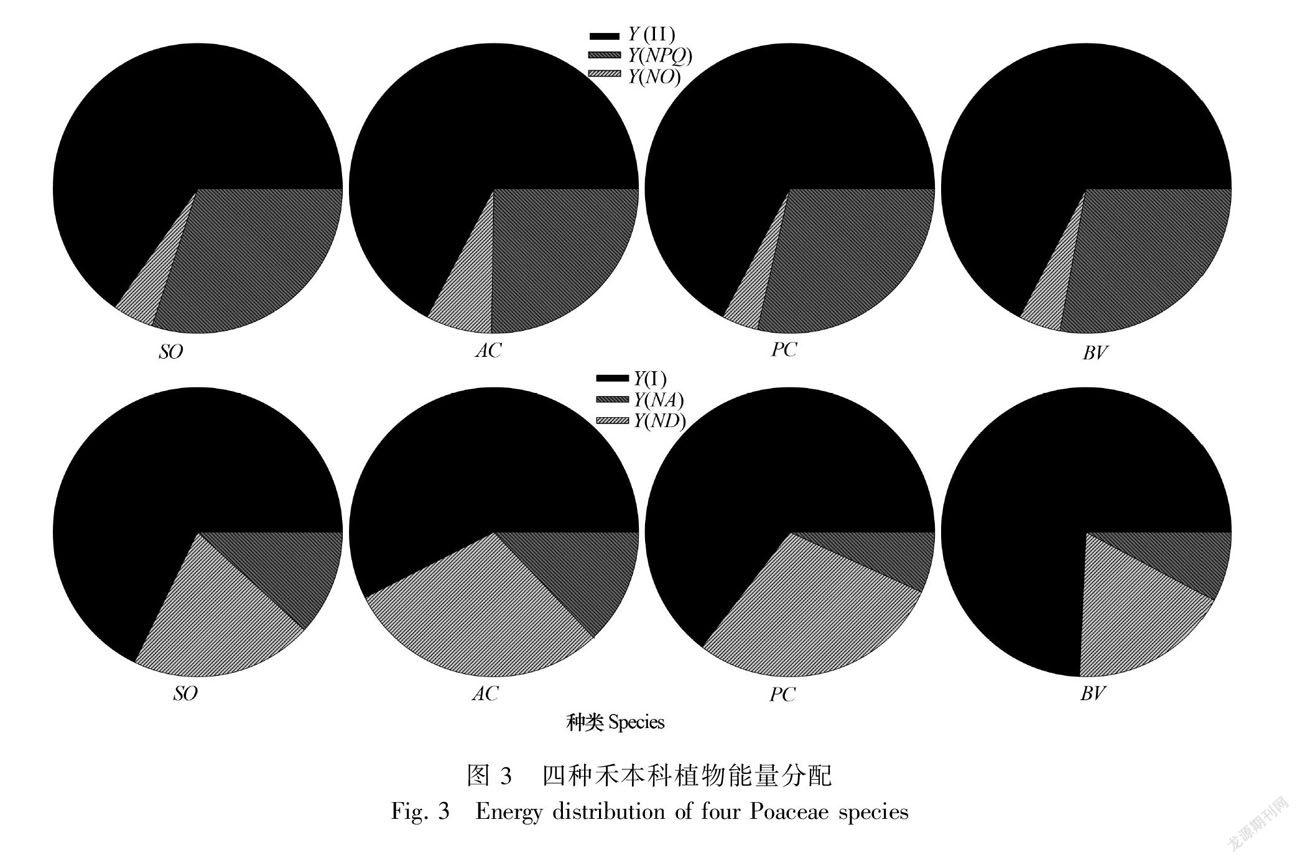
从四种植物PS Ⅱ的能量利用看,总体上它们均通过Y(Ⅱ)消耗大量光能,Y(NO) 耗能额比例最小。种间比较,地毯草Y(NO) 在总能量中占比显著高于其他三种植物,Y(NPQ)则明显低于其他三种植物(图3,上排)。
四种植物PS Ⅰ的能量使用总体是Y(I) 耗能额度最大,其中在佛肚竹中占比最高,在地毯草中占比最低,芦苇和甘蔗居中(图3,下排)。Y(NA)在地毯草和芦苇中占比明显大于甘蔗和佛肚竹。Y(ND) 能耗在芦苇中占比相对较低,其次是佛肚竹,甘蔗和地毯草中则相对较大。
2.3 四种禾本科植物光合气体交换能力
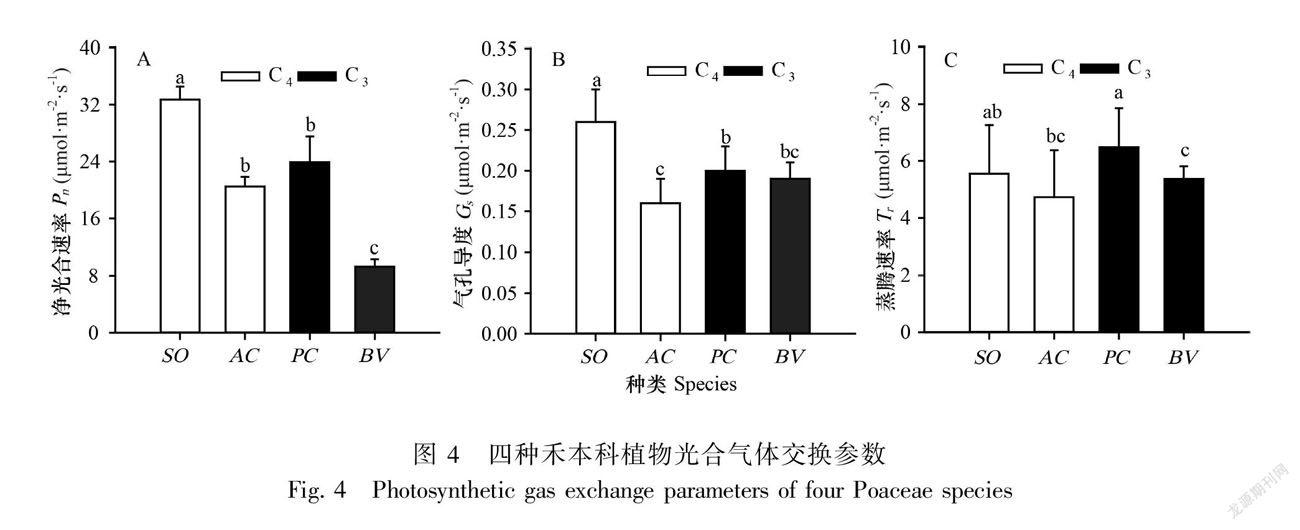
从Pn结果看,甘蔗最高,芦苇和地毯草居中,佛肚竹则最低(图4:A)。Gs方面,甘蔗同样最高,芦苇、佛肚竹和地毯草依次降低(图4:B)。与Pn和Gs不同,Tr在四种植物中大小依次为芦苇>甘蔗>地毯草>佛肚竹(图4:C)。
2.4 四种禾本科植物光响应曲线
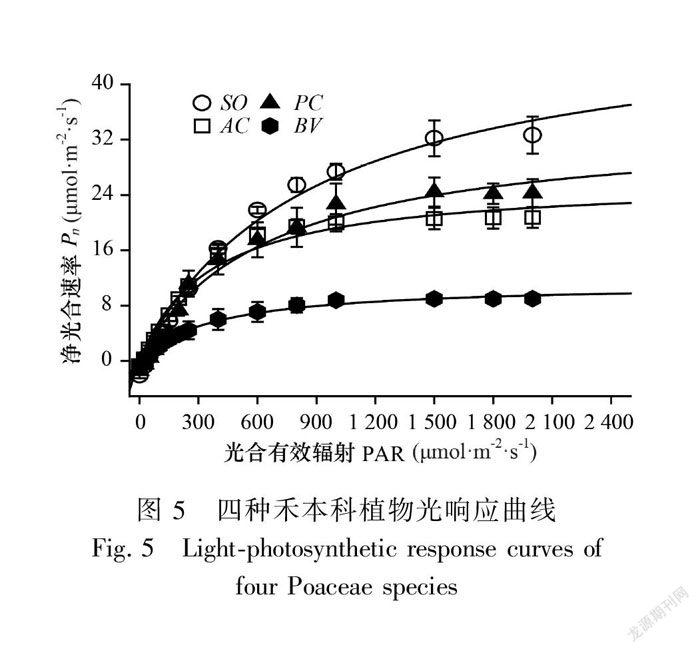
四种植物的光响应曲线如图5所示。甘蔗表现出典型C4植物的特征,即使在炎热天气条件下,光响应曲线也没有明显的饱和倾向。C3的芦苇也表现出部分C4植物的曲线特征,甚至比C4的地毯草更明显。佛肚竹明显不同于其他三种植物,光响应曲线是典型的C3植物特征。在低光区(<500 μmol·m-2·s-1),甘蔗、芦苇和地毯草Pn值比较接近,在中高光区(500~2 000 μmol·m-2·s-1)则逐渐显现出差异。与其他三种植物相比,无论在低光区还是高光区,佛肚竹Pn值均明显较低。
2.5 四种禾本科植物水分利用率、量子效率(Φi)及相关性
如图6所示,从种间对比看,芦苇和地毯草的WUEi较高,甘蔗其次,而佛肚竹则远低于三者。甘蔗和地毯草的WUE较高,其次是芦苇,佛肚竹同样远低于三者。地毯草的Φi较高,其次是甘蔗和芦苇,佛肚竹远低于前三者。
综合分析结果显示,四种植物炎夏伏天WUE和Φi总体上呈显著的線性正相关关系(图6:A,B)。
2.6 四种禾本科植物几个重要辅助光合参数
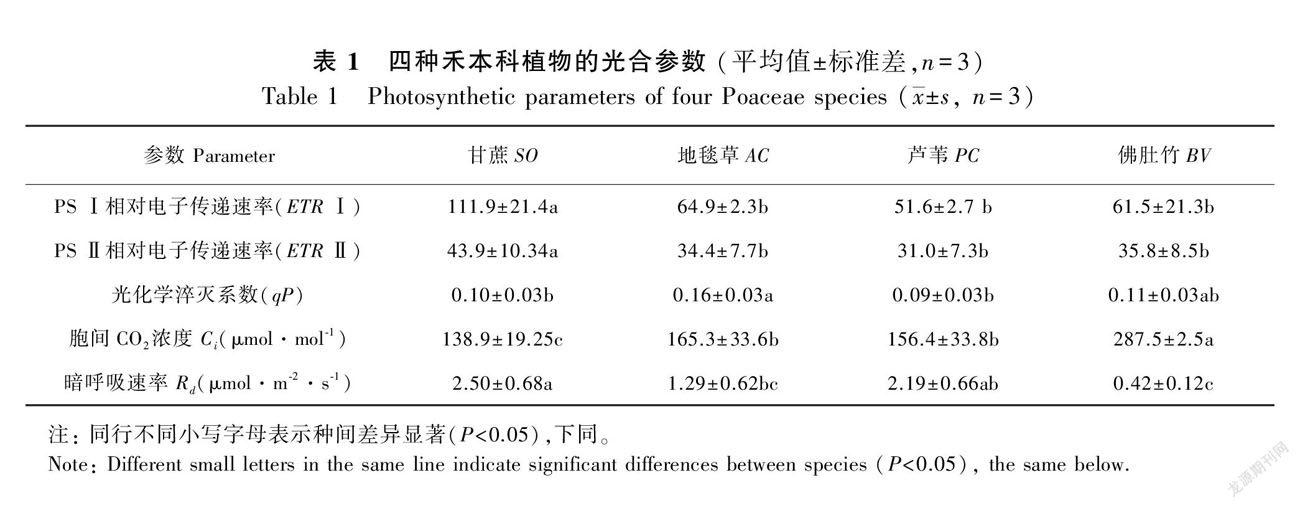
如表1所示,四种植物中甘蔗的ETR Ⅰ最高,地毯草和佛肚竹居次,最低是芦苇。甘蔗的ETR Ⅱ最高,其他三种植物无显著差异。地毯草的qP较高,其次是佛肚竹,芦苇和甘蔗则相对较低。佛肚竹的Ci最高,其次是地毯草和芦苇,甘蔗则较低。甘蔗的暗呼吸速率(Rd)最高,其次是芦苇和地毯草,佛肚竹最低。
2.7 四种禾本科植物光合色素含量
四种植物色素含量见表2,新黄质含量种间差异显著,佛肚竹最高,地毯草最低。叶黄质含量的种间差异显著,以芦苇最高,地毯草最低。佛肚竹的叶黄素循环库容显著高于芦苇,后者显著高于甘蔗,地毯草显著低于其他三种植物。甘蔗和芦苇的叶绿素总量较高,其次是佛肚竹,地毯草最低。总体上来看,地毯草光合相关色素含量显著低于其他三种植物(P<0.05)。
2.8 四种禾本科植物光合色素含量和光合性能的相关性
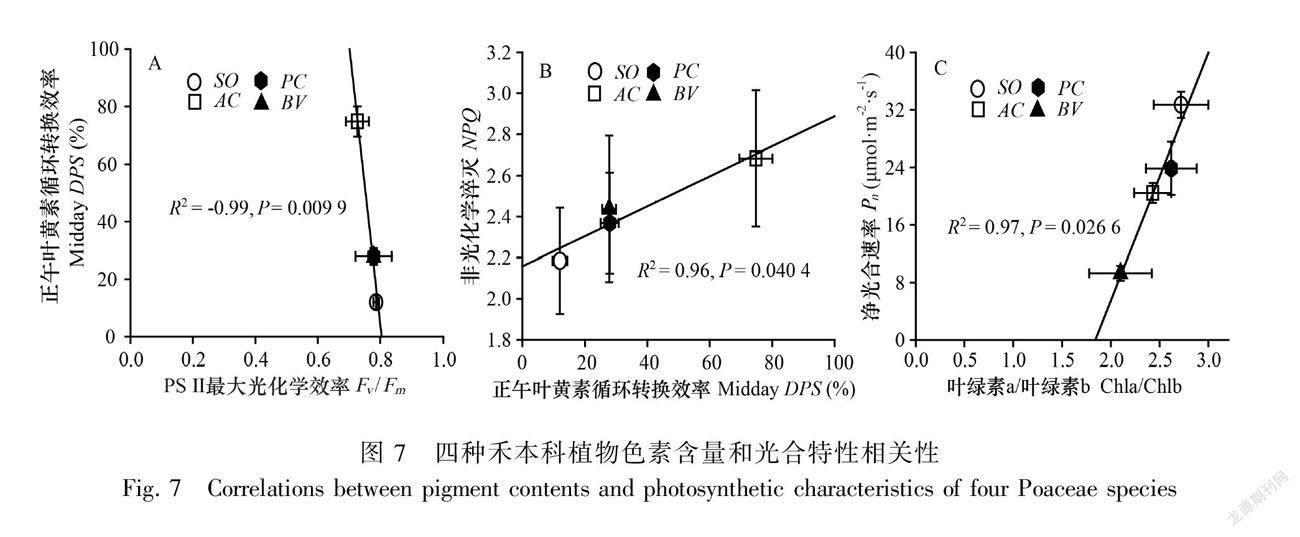
综合分析光合特性和色素含量的相关性,结果显示,Fv/Fm和正午DPS极显著负相关(图7:A),而DPS与NPQ显著正相关(图7:B),Chla/Chlb的比值与净光合速率显著正相关(图7:C)。
3 讨论
3.1 C4 型与C3型禾本科植物在南亚热带地区炎夏伏天光能利用策略及差异分析
总体上看,南亚热带四种禾本科植物的光合作用较好地适应了炎夏高温潮湿天气。虽然C4的地毯草有轻微光抑制,但作为地生植物已较强。
四种植物光能利用策略差异明显。从两种C4植物来看,它们同属于禾本科系统树中的PACMAD分支中的黍亚科(与虎尾草亚科并称该分支的两大骨干群系),过去该亚科在系统树中位于较高级分支,近来根据叶绿体全基因组的研究结果 (Cotton et al., 2015) 及辅助研究,将其调为PACMAD分支的基部群系(Soreng et al., 2015),在族、亚族和属的水平上,两种C4植物处于并列的单系群中。这两种C4植物最突出的光能利用策略是在高热天气下依然维持高Pn,从而消耗大量光能。C4途径的进化形成机制目前主要有两种学说。其一是环境学说,认为低CO2浓度和干旱是驱动C3途径进化为C4途径的根本动力,为适应低CO2浓度,C4植物进化出浓缩CO2的维管束“鞘泵”和高亲和CO2的磷酸烯醇式丙酮酸羧化酶(PEP羧化酶)(Christin et al., 2008;Christin & Osborne, 2014)。PACMAD分支和BOP分支虽然有共同的先祖,但只有前者进化出C4光合途径,其根本原因是环境(低CO2浓度、高温、干旱)选择压力不同,此外,环境压力下基因组中进化出一些编码酶的基因,可能促进了C4途径的形成(Griffiths et al., 2013)。其二是解剖学说,认为PACMAD分支先祖具有比BOP分支先祖更大的维管束鞘(BS)面积(前者中占比大于15%),更小的BS间距,在低CO2浓度和干旱胁迫发生时,PACMAD分支BS间距逐渐变小,而BS面积逐渐加大,最终进化出C4途径,BOP分支中BS面积减小是C4途径分化完成后才发生的(Christin et al., 2013)。近年来,该学说得到越来越多的支持(Lundgren et al., 2019)。无论是环境学说还是解剖学说,与C3植物相比,C4植物客观上进化出了更加亲和CO2的酶系和含叶绿体的“BS泵”, 叶绿体分布格局也出现了明显的变化。这种变化不但更适应低CO2浓度,客观上也促进了更高效的水分运输。基于此,有观点认为,C4途径演化的结果是同时优化了碳、水利用效率(Osborne & Sack, 2012)。本研究中,两种C4植物光合速率和水分利用效率均较高,支持上述观点。秦茜等(2015)比较7个具有不同抗旱能力的甘蔗品种发现,抗旱能力强的品种具有更小的脉间距、更大的BS面积和更高的Pn。由此可见,甘蔗中碳、水优化利用策略联系紧密,炎夏高光高温条件下这种关系再次得到体现。叶子飘等(2016)研究发现,C4玉米和高粱比C3的栾树和辣椒有更高的水分利用效率,这些结果同样也支持上述C4途径演化中碳、水利用协同优化的观点。此外,本研究还发现,C4植物的高WUE与高Φi紧密联系。Furbank & Hatch (1987)指出,强光高温条件下,C4植物BS会渗漏部分CO2到叶肉细胞中去,从而提高叶肉细胞的Φi。本研究中,两种C4植物高WUE极可能也是高碳同化力的一个客观反映,同时还提高了叶片的Φi,增强叶片弱光利用能力,综合光适应能力更强。
高温炎热天气条件下,C4地毯草叶绿素总含量明显低于其他三种植物,有助于减少叶片对强光的吸收,这是地生C4植物的一种光适应策略。和5月常温条件(常温下叶绿素总量为9.8±0.3 mg·g-1)相比较,C4甘蔗叶绿素总含量也降低,叶绿素a/b的比值也略低于常温条件(常温下叶绿素a/b值为3.1∶1)。色素含量和色素-蛋白复合体,尤其是捕光蛋白复合体有固定化学计量关系(Farquhar et al., 1982),色素含量的降低是叶绿体内部结构和功能调节的一种反映,有助于防止光氧化和光漂白(Powles, 1984; Niyogi et al., 2001)。本研究还发现,Chla/Chlb和Pn呈显著的正相关关系,表明Chla/Chlb比值的变化直接影响光合碳同化能力。
逆境胁迫下,通过维持高Pn消耗光能的策略在部分C3植物,如红树(朱俊杰和曹坤芳,2018)、冬小麦(Savitch et al., 2002)等植物上也有报道。本研究中,处于禾本科进化树PACMAD分支较高位置芦竹亚科的C3芦苇,和该分支基群系黍亚科的两种C4植物类似,也通过维持较高的Pn消耗大量光能,并且,芦苇同样具有较高的WUE和Φi。龚春梅等(2007)发现,在干旱胁迫下部分芦苇会出现由C3向C4途径的转化,这种光合途径转化现象在C4植物不同亚型间也有报道(Wang et al., 2014),被子植物C4途徑的形成至少经历了64次独立进化(Hibberd & Quick, 2002),禾本科植物中则至少有22~24次独立进化(GPWG Ⅱ et al., 2012),逆境下光合途径转化的现象与此有关。
高温湿热天气下,芦苇利用类胡萝卜素防御光抑制也是一种光能利用策略。相比于其他三种植物,芦苇叶黄质和新黄质含量均较高,叶黄素循环库容也较高。这些类胡萝卜素同处于一个代谢体系中,叶黄素循环色素可以转换为新黄质,其前体物质也可以转换为叶黄质(Lu & Li, 2008),类胡萝卜素总体较高表明该代谢途径活跃。叶黄素循环的光保护作用机制有玉米黄质直接淬灭激发态叶绿素假说(Chow, 1994)和PS Ⅱ色素捕光蛋白复合体聚集利于叶绿素荧光非淬灭假说(Horton et al., 1996)。从叶黄素循环色素中午DPS看,芦苇处于中等水平,正午DPS和 NPQ显著线性正相关,表明叶黄素循环实质上起到了光保护作用,支持色素捕光蛋白复合体聚集淬灭假说。
佛肚竹位于禾本科系统树BOP分支的较高位,与两种C4植物相比,在炎夏伏天固碳能力及水分利用率均较低,量子效率也较低,但其叶黄素循环色素库容最高。其光能利用策略最明显的特征是借助类胡萝卜素与NPQ偶联方式耗散多余光能。孙化雨等(2015)和娄永峰(2016)在毛竹中也证实了叶黄素循环偶联NPQ的光抑制防御策略。
3.2 炎夏伏天光能利用策略与生态分布和生活型选择的关系分析
本研究中,南亚热带地区两种C4禾本科植物是旱生生态型,两种C3植物是湿生生态型。从演化角度分析,两种C4植物特殊的叶片结构和酶系使它们在适应旱生生境方面优势明显。野生甘蔗和栽培甘蔗几乎遍布整个南亚带气候区,是典型的拓荒先锋植物。C3芦苇在南亚热带地区分布范围明显小于甘蔗,主要分布在河滩、水塘等潮湿环境,部分芦苇也能分布到较干旱的滩涂和干涸的河滩,其较强的碳同化能力及通过调配类胡萝卜素耗散光能的机制起了保障作用。至于这些旱生地段的芦苇是否演化出C4光合途径有待进一步确认。佛肚竹光合能力较弱,水分利用效率低,主要通过色素调配应对高温强光胁迫,而色素底物主要来自光合产物,这种生理特性限制了它们的分布范围。和前述三种植物相比,其分布范围更狭小,主要散布于相对湿度较大的水塘、沟渠边上,并且营丛生生活。
从生活型分析,地毯草是典型的地面芽多年生草本植物,其他三种植物是高位芽多年生草本植物。在自然条件下,地面芽植物很难保证其上方没有其他植物遮盖,因此,其生态策略偏向于阴生。虽然地毯草进化出C4光合途径,但其光能利用策略相对保守,体现在:第一,其PS Ⅱ总能量耗散中Y(NO)比例几乎是其他三种植物的2倍,PS I总能量耗散中Y(ND)+Y(NA)的和也明显高于其他三种植物;第二,Gs、Tr相对较低;第三,类胡萝卜素和叶绿素含量均显著低于其他三植物,在高温炎热环境下除了维持一定的Pn外,也通过大幅提高DPS来加强热耗散。研究小组观察到地毯草在低温条件下合成大量的花青素(待发表数据)作为临时光保护物质,其他三种植物则较少有这种次生代谢,也从一个侧面反映出其光能利用机制的保守性。
4 结论
总之,南亚热带地区炎夏伏天高热天气条件下, C4甘蔗通过高光合速率消耗大量光能;C4地毯草通过较高碳同化并结合较高DPS加强热耗散;C3芦苇既有类似C4植物的高固碳和高水分利用效率,也通过类胡萝卜素耦合NPQ耗散部分光能;C3的佛肚竹主要依靠类胡萝卜素耦合NPQ耗散多余光能。这些植物光能利用策略与它们的生态分布和生活型选择密切关联。
参考文献:
ACEBRON K, MATSUBARA S, JEDMOWSKI C, et al., 2020. Diurnal dynamics of nonphotochemical quenching in Arabidopsis npq mutants assessed by solar-induced fluorescence and reflectance measurements in the field[J]. New Phytol, 229(4): 2104-2119.
ASADA K, 1999. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons[J]. Ann Rev Plant Physiol, 50(1): 601-639.
CHEN MD, ZHU HS, WANG B, et al., 2016. Analysis of xanthophyll cycle components in strawberry leaves by ultra-high performance liquid chromatography[J]. Chin J Trop Crops, 37(9): 1841-1847. [陳敏氡,朱海生,王彬,等, 2016. 草莓叶叶黄素循环组分超高效液相色谱分析[J]. 热带作物学报, 37(9): 1841-1847.]
CHOW WS, 1994. Photoprotection and photoinhibition[M]// BITTAR EE. Advances in molecular and cell biology. Greenwich: Elsevier: 151-196.
CHRISTIN PA, BESNARD G, SAMARITANI E, et al., 2008. Oligocene CO2 decline promoted C4 photosynthesis in grasses[J]. Curr Biol, 18(1): 37-43.
CHRISTIN PA, OSBORNE CP, 2014. The evolutionary ecology of C4 plants[J]. New Phytol, 204(4): 765-781.
CHRISTIN PA, OSBORNE CP, CHATELET DS, et al., 2013. Anatomical enablers and the evolution of C4 photosynthesis in grasses[J]. Proc Natl Acad Sci USA, 110(4): 1381-1386.
COTTON JL, WYSOCKI WP, CLARK LG, et al., 2015. Resolving deep relationships of PACMAD grasses: A phylogenomic approach[J]. BMC Plant Biol, 15(1): 178.
EDWARDS EJ, OSBORNE CP, STRMBERG CAE, et al., 2010. The origins of C4 grasslands: Integrating evolutionary and ecosystem science[J]. Science, 328(5978): 587-591.
FURBANK RT, HATCH MD, 1987. The size and composition of the inorganic carbon pool in bundle-sheath cells[J]. Plant Physiol, 85(4): 958-964.
GONG CM, 2007. Studies on the adaptive change mechanism of photosynthetic carbon assimilation pathway in plants under water gradient in arid regions[D]. Lanzhou: Lanzhou University: 32-35. [龚春梅, 2007. 干旱地区水分梯度下植物光合碳同化途径适应性变化机制研究[D]. 兰州: 兰州大学: 32-35.]
GRASS PHYLOGENY WORKING GROUP I, BARKER NP, CLARK LG, et al., 2001. Phylogeny and subfamilial classification of the grasses (Poaceae)[J]. Ann Mo Bot Gard, 88(3): 373-457.
GRASS PHYLOGENY WORKING GROUP II, ALISCIONI S, BELL HL, et al., 2012. New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins[J]. New Phytol, 193(2): 304-312.
GRIFFITHS H, WELLER G, TOY LFM, et al., 2013. You’re so vein: Bundle sheath physiology, phylogeny and evolution in C3 and C4 plants[J]. Plant Cell Environ, 36(2): 249-261.
HIBBERD JM, QUICK WP, 2002. Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants[J]. Nature, 415(6870): 451-454.
HOLT NE, ZIGMANTAS D, VALKUNASL, et al., 2005. Carotenoid cation formation and the regulation of photosynthetic light harvesting[J]. Science, 307(5708): 433-436.
HORTON P, RUBAN AV, WALTERS RG, 1996. Regulation of light harvesting in green plants[J]. Ann Rev Plant Physiol, 47(1): 655-684.
LI XP, BJRKMAN O, SHIH C, et al., 2000. A pigment-binding protein essential for regulation of photosynthetic light harvesting[J]. Nature, 403(6768): 391-395.
LLOYD J, FARQUHAR GD, 1994. 13C Discrimination during CO assimilation by the terrestrial biosphere[J]. Oecologia, 99(3-4): 201-215.
LOU YF, 2016. Study on light protection and related gene function of Phyllostachys pubescens [D]. Beijing: Chinese Academy of Forestry Sciences: 97-108. [婁永峰, 2016. 毛竹光保护及相关基因功能研究[D]. 北京: 中国林业科学研究院: 97-108.]
LU S, LI L, 2008. Carotenoid metabolism: biosynthesis, regulation, and beyond[J]. J Integr Plant Biol, 50(7): 778-785.
LUNDGREN MR, DUNNING LT, OLOFSSON JK, et al., 2019. C4 anatomy can evolve via a single developmental change[J]. Ecol Lett, 22(2): 302-312.
MATSUBARA S, CHOW WS, 2004. Populations of photoinactivated photosystem II characterized by chlorophyll fluorescence lifetime in vivo[J]. Proc Natl Acad Sci USA, 101(52):18234-18239.
NIXON PJ, RICH PR, 2007. Chlororespiratory pathways and their physiological significance[M]// WISE RR, HOOBER JK. The structure and function of plastids: Advances in photosynthesis and respiration (23). Dordrecht: Springer:237-251.
NIYOGI KK, BJRKMAN O, GROSSMAN AR, 1998. Arabidopsis mutants define a central role for the xanthophyll cycle in the regulation of photosynthetic energy conversion[J]. Plant Cell, 10(7): 1121-1134.
NIYOGI KK, SHIH C, CHOW WS, et al., 2001. Photoprotection in a zeaxanthin- and lutein-deficient double mutant of Arabidopsis[J]. Photosynth Res, 67(1-2): 139-145.
ORT DR, BAKER NR, 2002. A photoprotective role for O2 as an alternative electron sink in photosynthesis?[J]. Curr Opin Plant Biol, 5(3): 193-198.
POWLES SB, 1984. Photoinhibition of photosynthesis induced by visible light[J]. Ann Rev Plant Biol, 35(1): 15-44.
QIN X, ZHU JJ, GUAN XY, et al., 2017. The correlations of leaf anatomical characteristics with photosynthetic capacity and drought tolerance in seven sugarcane cultivars[J]. J Plant Physiol, 53(4): 705-712.[秦茜, 朱俊杰, 关心怡, 等, 2017. 七个甘蔗品种叶片解剖结构特征与光合能力和耐旱性的关联[J]. 植物生理学报, 53(4): 705-712.]
RAVEN JA, 2020. Chloride involvement in the synthesis, functioning and repair of the photosynthetic apparatus in vivo[J]. New Phytol, 227(2): 334-342.
SAGE RF, 2016. A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and hall of fame[J]. J Exp Bot, 67(14): 4039-4056.
SAVITCH LV, LEONARDOS ED, KROLM, et al., 2002. Two different strategies for light utilization in photosynthesis in relation to growth and cold acclimation[J]. Plant Cell Environ, 25(6): 761-771.
SORENG RJ, PETERSON PM, ROMASCHENKO K, et al., 2015. A worldwide phylogenetic classification of the Poaceae (Gramineae)[J]. J Syst Evol, 53(2): 117-137.
SUN HY, CHEN Y, ZHAO HS, et al., 2015. Molecular characteristics and functional analysis of β-carotene hydroxylase gene from Phyllostachys edulis[J]. Sci Silv Sin, 51(10): 53-58.[孙化雨, 陈颖, 赵韩生, 等, 2015. 毛竹β-胡萝卜素羟化酶基因的分子特征及其功能[J]. 林业科学, 51(10): 53-58.]
YE ZP, YANG XL, KANG HJ, 2016. Comparison of light-use and water-use efficiency for C3 and C4 species[J]. Acta Agric Zhejiang, 28(11): 1867-1873.[叶子飘, 杨小龙, 康华靖, 2016. C3和C4植物光能利用效率和水分利用效率的比较研究[J]. 浙江农业学报, 28(11): 1867-1873.]
YE ZP, YU Q, 2008. A coupled model of stomatal conductance and photosynthesis for winter wheat[J]. Photosynthetica, 46(4): 637-640.
ZHU JJ, 2007. Physiological and ecological mechanism of plant light protection and antioxidant protection in Yuanjiang dry and hot valley[D]. Xishuangbanna: Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences: 87-93. [朱俊杰, 2007. 元江干热河谷植物光保护和抗氧化保护生理生态机制[D]. 西双版纳: 中国科学院西双版纳热带植物园: 87-93.]
ZHU JJ, CAO KF, 2018. Some special phenomena of photosynthesis in mangrove plants[G]. Kunming: Collection of abstracts from papers presented at the 85th Annual Meeting of the National Botany Society (1993-2018). [朱俊杰, 曹坤芳, 2018. 红树植物光合作用的一些特殊现象[G]. 昆明: 中国植物学会八十五周年学术年会论文摘要汇编(1993-2018).]
(责任编辑 周翠鸣)
收稿日期:2021-03-27
基金项目:国家自然科学基金(31460102)[Supported by National Natural Science Foundation of China (31460102)]。
第一作者: 張雅芳(1996-),硕士研究生,研究方向为植物光合生理生态,(E-mail)314794036@qq.com。
通信作者:朱俊杰,博士,副教授,研究方向为植物光合生理生态,(E-mail)20150067@gxu.edu.cn。
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
今日农业(2022年13期)2022-09-15
环球时报(2022-06-20)2022-06-20
廉政瞭望·下半月(2021年8期)2021-09-18
现代农业科技(2018年4期)2018-03-23
决策(2017年8期)2017-09-06
