
基于高通量测序的魔芋根际土壤真菌多样性分析
2022-05-28 09:09王佰涛杨文玲李珊珊刘德海
西南农业学报 2022年4期
王佰涛, 杨文玲, 雷 高, 李珊珊, 刘德海
(河南省科学院生物研究所有限责任公司, 郑州 450008)
【研究意义】魔芋(Amorphophalluskonjac, K.Koch),别名蛇玉米、鬼芋,属南天星科魔芋属,产地主要在中国、日本等东南亚国家[1]。作为世界卫生组织公布的“十大保健食品”之一,魔芋本身营养丰富,其作为一些地区的重要食物来源种植历史悠久,当前在食品、医疗、畜牧等领域应用较多[2-4]。魔芋种植过程中对生长环境要求较高,不仅需要合适的海拔和湿度,魔芋根际微生物平衡对魔芋的生长也非常重要,当魔芋根际微生物失衡时更易导致病害的发生[5-6],目前对魔芋根际土壤细菌多样性研究较多,而对真菌群落多样性研究较少。因此,了解魔芋根际真菌群落多样性不仅有利于了解魔芋的生长情况和病害情况,还能促进魔芋行业的发展[7]。【前人研究进展】魔芋的主要成分是葡甘露聚糖(Konjac Glucomannan, KGM)[8],葡甘露聚糖主要是降解成为葡甘露低聚糖进行应用[9]。目前存在一些物理化学方法[10],但生物方法利用微生物发酵降解,具有绿色、高效等优势好[11-14],分析根际土壤真菌群落多样性有助于寻找相关微生物,而目前关于魔芋根际土壤细菌多样性分析的研究内容较多[15],对魔芋根际土壤真菌群落研究内容较少。【本研究切入点】魔芋根际真菌群落的生态功能对于魔芋生长非常重要,其不仅能够促进魔芋生长,还具备降解农药及重金属污染、防治病害等功能[16-17]。微生物测序技术优势明显,其快速、高效、精确的特点在微生物多样性分析中应用广泛[18]。如图1所示,本研究通过从不同地区采集魔芋根际土壤样品,利用Illumina MiSeq高通量测序对魔芋根际土壤真菌群落进行了多样性分析。【拟解决的关键问题】对不同地区的根际土壤真菌多样性进行了对比,详细分析了不同地区真菌群落在门、纲、属不同水平的优势菌属和丰度高低,以期为魔芋的产业化发展提供帮助,为实现高效制备葡甘露低聚糖奠定基础。

图1 魔芋根际土壤真菌群落多样性分析流程
1 材料与方法
1.1 试验地概况
试验样品地点为3处(表1)。在每个取样地分别随机选择3株魔芋进行取样,取样时,除去表层土壤,在0~20 cm深度范围内采集魔芋根际土壤。将采集的魔芋根际土壤样品快速保存至低温保藏箱中,无菌条件研磨后用于实验。

表1 采样信息统计
1.2 实验设计
试验设置3组样品,每个样品设置3个重复,分别是昆明样品(A):A1、A2、A3;南平样品(B):B1、B2、B3;南阳样品(C):C1、C2、C3。
1.3 测定项目及方法
1.3.1 基因组提取和PCR扩增 根据 DNA提取试剂盒说明书进行微生物群落总 DNA 提取,使用1%的琼脂糖凝胶电泳检测DNA的提取质量,使用NanoDrop2000测定DNA 浓度和纯度;使用ITS1F(5’-CTTGGTCATTTAGAGGAAGTAA-3’)和ITS2R(5’-GCTGCGTTCTTCATCGATGC-3’)对核糖体核糖核酸(ribosomal ribonucleic acid, rRNA)基因ITS区进行 PCR 扩增。
1.3.2 Illumina Miseq 测序 将同一样品的PCR产物混合后使用2%琼脂糖凝胶回收PCR产物,利用DNA凝胶回收试剂盒进行回收产物纯化,1%琼脂糖凝胶电泳检测,并用微型荧光计对回收产物进行检测定量。使用DNA建库试剂盒进行建库: ①接头链接;②使用磁珠筛选去除接头自连片段;③利用PCR扩`增进行文库模板的富集;④磁珠回收PCR产物得到最终的文库。利用Illumina公司的Miseq PE300平台进行测序(上海美吉生物医药科技有限公司)。原始数据上传至NCBI 序列读取(Sequence read archive,SRA)数据库。
1.4 数据优化与聚类分析
使用Trimmomatic软件原始测序序列进行质控,使用FLASH软件进行拼接:使用UPARSE软件操作分类单元(Operational taxonomic unit,OTU)聚类,根据 97%的相似度对序列进行OTU聚类,使用UCHIME 软件剔除嵌合体,利用RDP classifier 软件对每条序列进行物种分类注释。
2 结果与分析
2.1 三地区真菌序列统计和多样性分析
利用Illumina Miseq 测序之后分别获得有效序列数为A1:73 192、A2:69 003、A3:71 663、B1:72 179、B2:68 728、B3:70 393、C1:71 077、C2:73 958、C3:67 715。对样品序列数进行抽平处理,抽平后序列数为67 715。在序列相似性97%水平进行序列划分OTU,9个样品共得到3770个OTUs。对3组样品进行Alpha多样性分析,Shannon指数值越高,Simpson指数值越低,说明群落多样性越高,Chao1指数或ACE指数越大,说明群落丰富度越高[19]。如表2所示,不同地区真菌群落多样性不同,比较3组样品的多样性指数、丰富度指数和均匀度指数,C样品的香农指数较高,辛普森指数较低,说明C样品的真菌群落多样性较高。C样品的Chao1指数和ACE指数对比A样品和B样品均较高,说明C样品真菌群落丰富度较高。总的来说,无论是丰富度还是多样性,C样品均较高。覆盖率越接近于1,说明测序深度越合理,测序深度已经基本覆盖到样品中的所有物种。由表可知A样品、B样品、C样品覆盖率均>99%,表明取样合理,测序结果能够真实反映昆明、南平、南阳3地的真菌群落。如图2所示,对3组数据OTU水平进行显著性差异分析,结果显示C样品的香农指数与A、B 样品存在显著性差异。通过3组样品对比分析发现,南阳样品为半年生魔芋植株,其在3组样品中真菌群落多样性最高,推测在魔芋种植半年以后逐渐建立微生态平衡,真菌群落多样性达到最大,南平样品为3年生魔芋植株,真菌群落多样性下降,说明随着年限的增长优势菌属的相对丰度逐渐增长,导致整体真菌群落多样性有所降低,而海拔高度和经纬度对魔芋根际土壤真菌群落多样性影响较小。
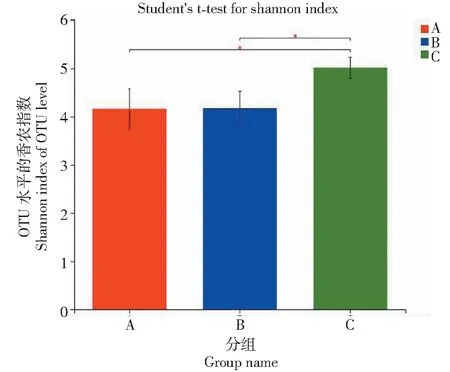
*代表显著相关(P<0.05)
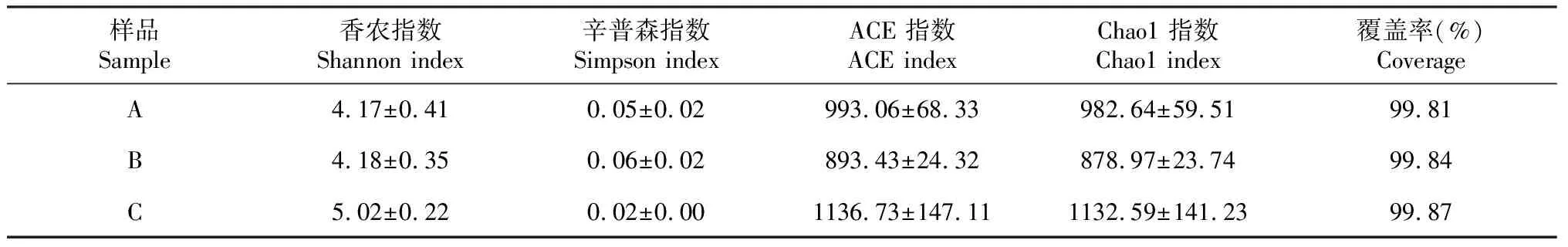
表2 不同地区样品真菌群落多样性分析
稀释曲线能够反映采集样品测序结果的覆盖度和真菌群落的测序深度[20],如图3所示,3组样品序列数达到4000 OTU水平时变化较大,随后随着测序数量的增加,稀释曲线趋于平稳,曲线变化趋势近似一条直线,更多的取样只会产生少量新的OTU,说明本次3组样品取样测序数据合理。
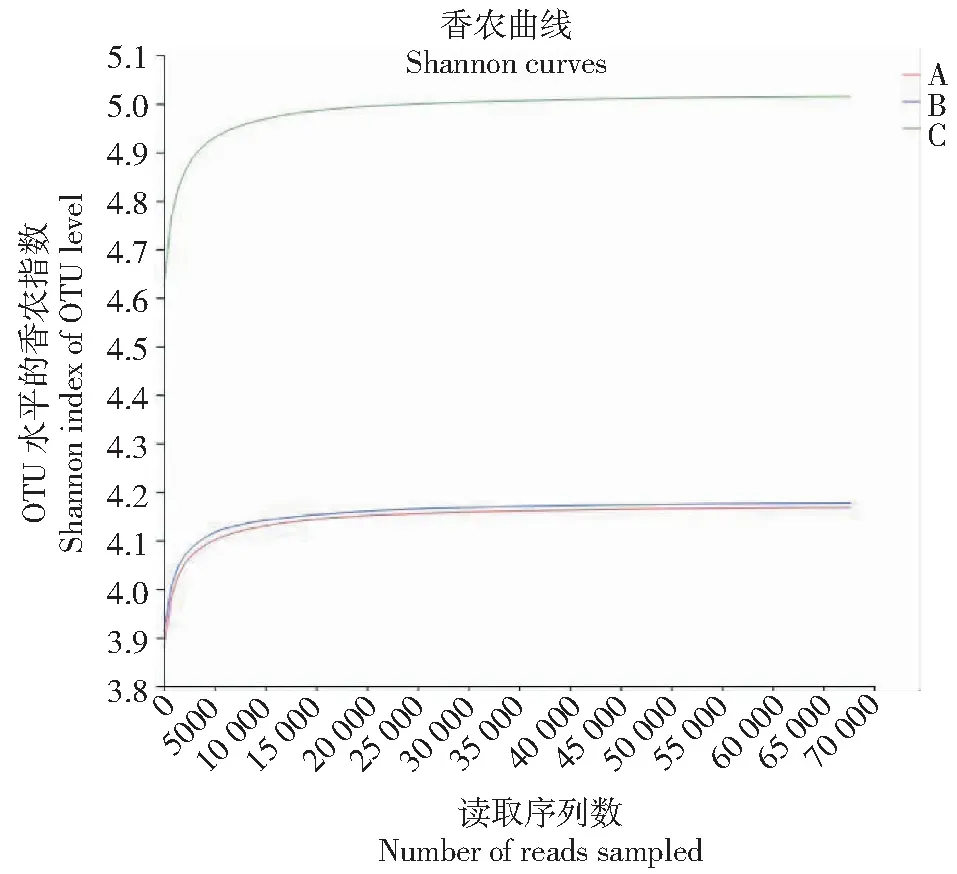
图3 基于香农指数不同地区样品的稀释曲线
2.2 真菌群落结构组成分析
2.2.1 昆明地区样品真菌菌落结构组成 A组样品真菌属于16个真菌门,55个纲,117个目,246个科,489个属。在门水平,有4个真菌门的丰度≥1%,是A组样品的优势菌群,其中子囊菌门(Ascomycota)的丰度为55.24%,担子菌门(Basidiomycota)的丰度为21.49%,被孢霉菌门(Mortierellomycota)的丰度为20.18%,其余12个门均低于1%,共占1.25%(图4)。
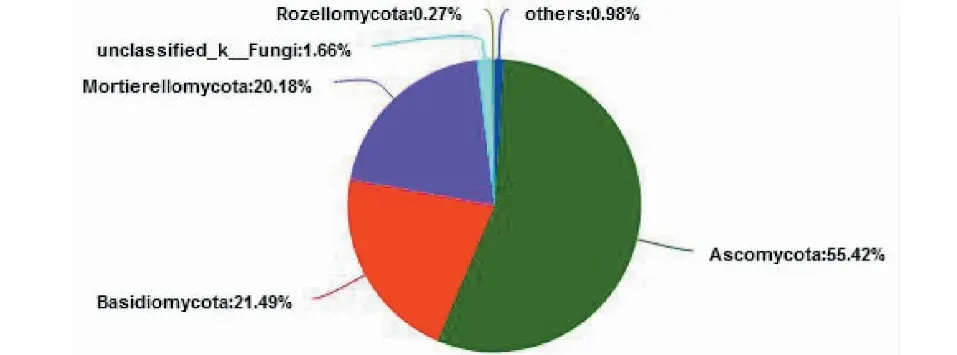
图4 A样品门水平真菌群落组成
如表3所示,在纲水平属于55个纲,相对丰度≥1%的纲有7个,其中子囊菌门的粪壳菌纲(Sordariomycetes,32.29%)丰度最高,被孢霉菌门的被孢霉纲(Mortierellomycetes,20.13%)次之,其他还有担子菌门的银耳纲(Tremellomycetes)等优势菌群。
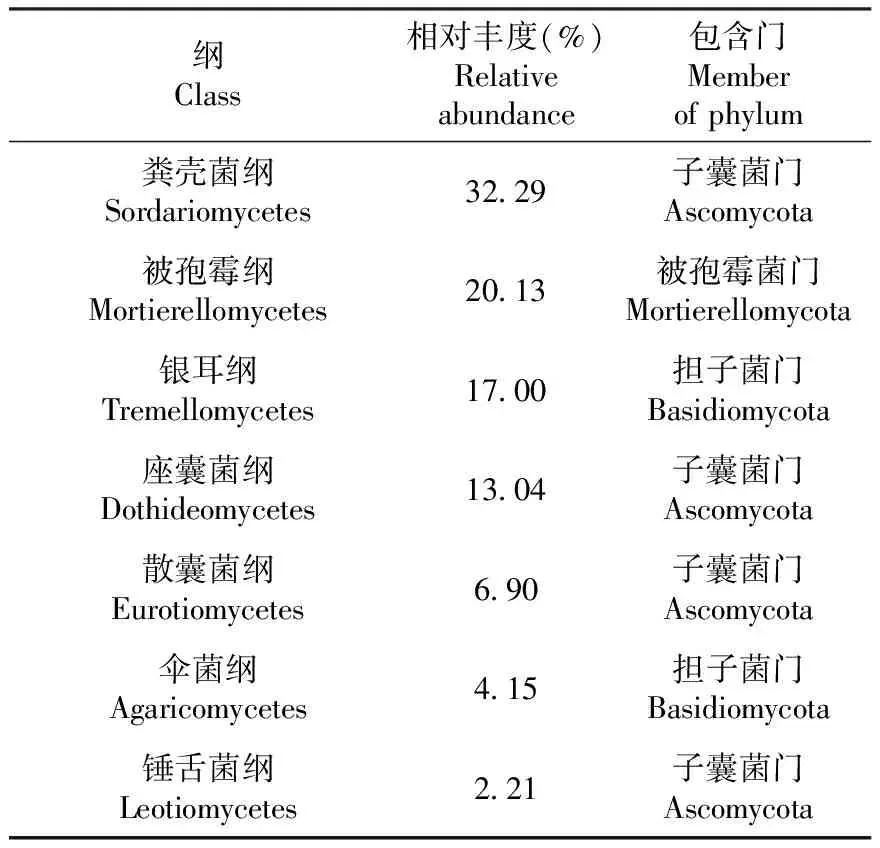
表3 A样品纲水平真菌组成及相对丰度
昆明地区样品真菌群落在属水平上共有489个属,其中有13个真菌属的丰度≥1%,属于优势菌属,主要包含被孢霉属(Mortierella)20.13%、赤霉属(Gibberella)6.07%、截盘多孢霉属(Truncatella)3.91%、镰刀霉属(Fusarium)3.25%、青霉属(Penicillium)2.57%和硬皮地星属(Astraeus)2.22%等(图5)。

图5 A样品属水平真菌群落组成
2.2.2 南平地区样品真菌菌落结构组成 B组样品真菌属于12个真菌门,38个纲,88个目,189个科,361个属。在门水平,有4个真菌门的丰度≥1%,是B组样品的优势菌群,其中子囊菌门(Ascomycota)的丰度为81.71%,担子菌门(Basidiomycota)的丰度为13.00%,被孢霉菌门(Mortierellomycota)的丰度为2.83%,未分类门为1.24%,其余8个门均低于1%,共占1.22%(图6)。

图6 B样品门水平真菌群落组成
如表4所示,在纲水平属于38个纲,相对丰度≥1%的纲有8个,其中子囊菌门的粪壳菌纲(Sordariomycetes,66.15%)丰度最高,担子菌门的伞菌纲(Agaricomycetes,8.05%)次之,其他还有子囊菌门的座囊菌纲(Dothideomycetes)等优势菌群。

表4 B样品纲水平真菌组成及相对丰度
南平地区样品真菌群落在属水平上共有361个属,有22个真菌属的丰度≥1%,属于优势菌属,主要包含赤霉属(Gibberella,17.77%)、亡革菌属(Thanatephorus,7.79%)、隐囊菌属(Aphanoascus,4.22%)、节担菌属(Wallemia,3.96%)、枝孢霉属(Cladosporium,3.50%)、镰刀霉属(Fusarium,2.90%)、绿僵菌属(Metarhizium,2.87%)、被孢霉属(Mortierella,2.72%)等(图7)。
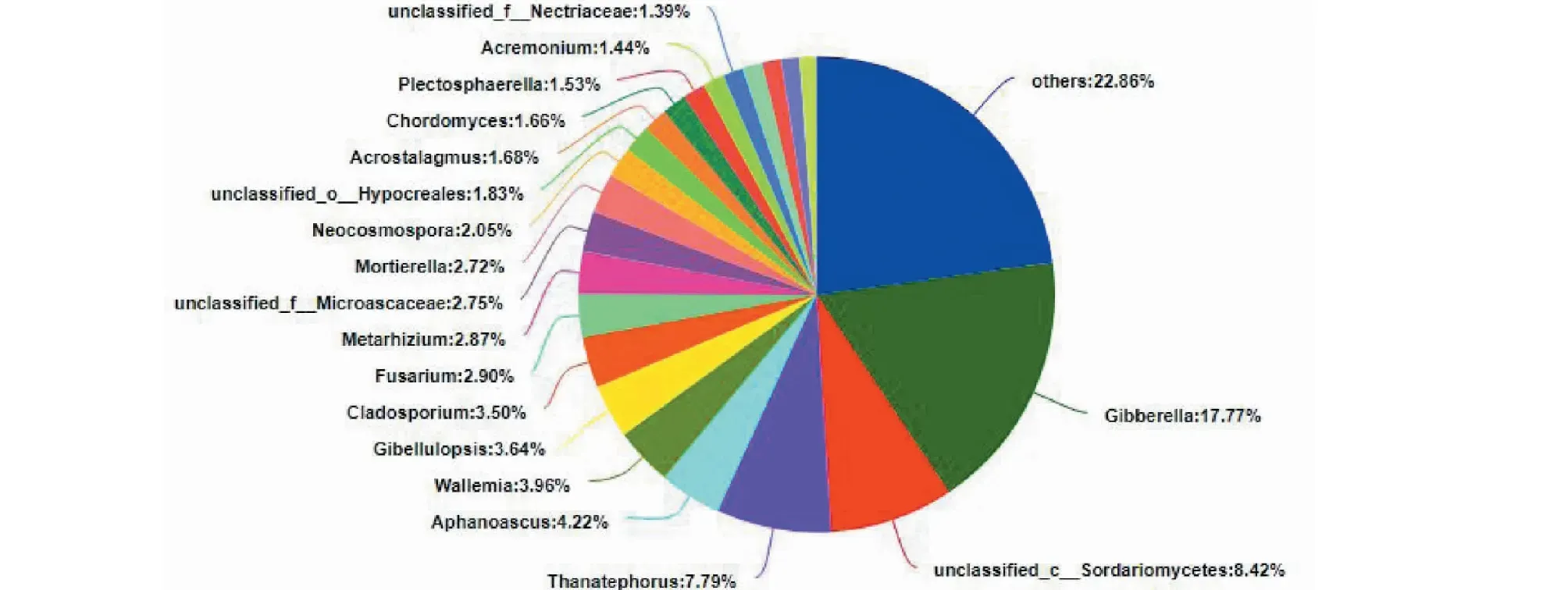
图7 B样品属水平真菌群落组成
2.2.3 南阳地区样品真菌菌落结构组成 C组样品真菌属于16个真菌门,48个纲,109个目,256个科,540个属。在门水平,有5个真菌门的丰度≥1%,是C组样品的优势菌群,其中子囊菌门(Ascomycota)的丰度为67.72%,担子菌门(Basidiomycota)的丰度为13.36%,被孢霉菌门(Mortierellomycota)的丰度为5.27%,罗兹菌门(Rozellomycota)的丰度为6.88%,未分类门为5.61%,其余11个门均低于1%,共占1.16%(图8)。

图8 C样品门水平真菌群落组成
如表5所示,在纲水平属于48个纲,相对丰度≥1%的纲有11个,其中子囊菌门的粪壳菌纲(Sordariomycetes,41.20%)丰度最高,子囊菌门的座囊菌纲(Dothideomycetes,13.35%)次之,其他还有担子菌门的银耳纲(Tremellomycetes)等优势菌群。
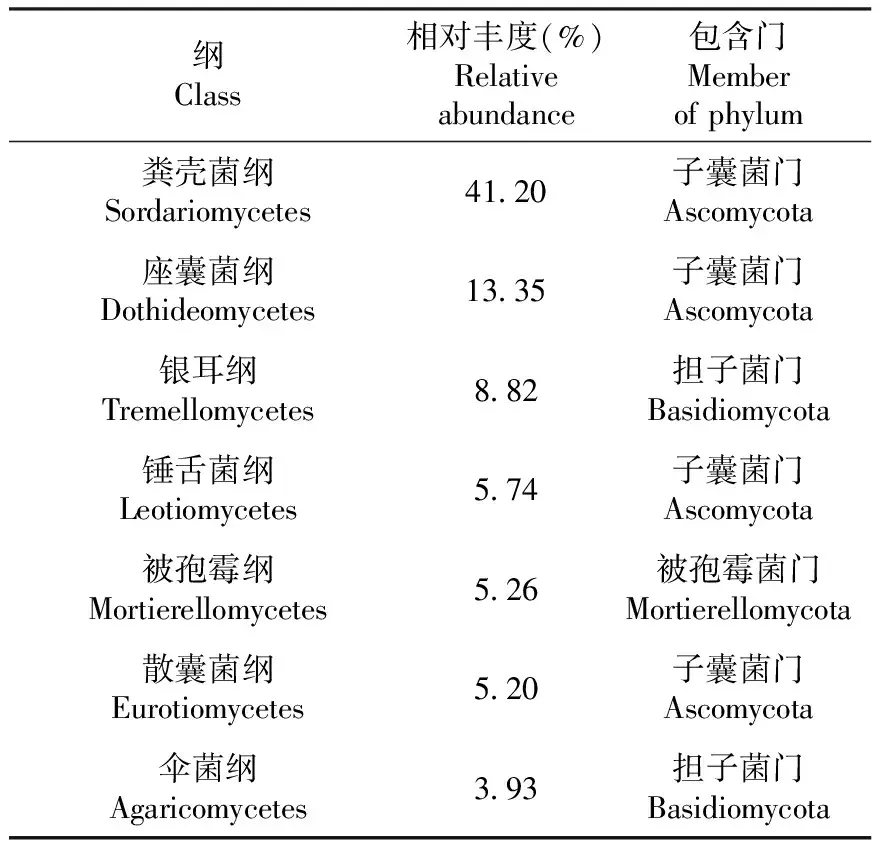
表5 C样品纲水平真菌组成及相对丰度
如图9所示,南阳地区样品真菌群落的丰富度和均匀度均较高,在属水平上共有540个属,有25个真菌属的丰度≥1%,属于优势菌属,主要包含Paraboeremia属(6.46%)、被孢霉属(Mortierella,5.26%、)新赤壳属(Neocosmospora,3.68%)、木霉属(Trichoderma,3.60%)、Solicoccozyma属(3.40%)、青霉属(Penicillium,2.99%)、镰刀霉属(Fusarium,2.60%)、Tausonia属(2.49%)、膝梗孢属(Gonytrichum,2.39%)、支孢霉属(Aremonium,2.21%)、绿僵菌属(Metarhizium,1.18%)等。其中木霉属真菌在我国工业、农业及环境修复方面都占据举足轻重的地位[21],木霉属真菌的生物降解转化及产纤维素酶在工业和农业上应用较多[22],并且通过研究发现利用木霉属真菌进行重金属污染修复方面也能表现不错的效果[23]。昆明样品中的优势菌属主要是被孢霉属(Mortierella)、截盘多孢霉属(Truncatella)、青霉属(Penicillium)和硬皮地星属(Astraeus,2.22%)等。其中,青霉属真菌为较常见且应用范围较广的真菌属,其在生物转化、医学及环保等方面应用较多[24],在自然界青霉菌属能够参与多种天然产物降解,也能产生多种蛋白酶、木质素酶等多种生物酶[25],还能在多环芳烃污染物处理方面发挥重要作用[26]。此外,青霉菌属的许多次级代谢产物在抗生素领域应用历史悠久[27],近年来其在癌症治疗方面的应用也有了新的突破[28]。
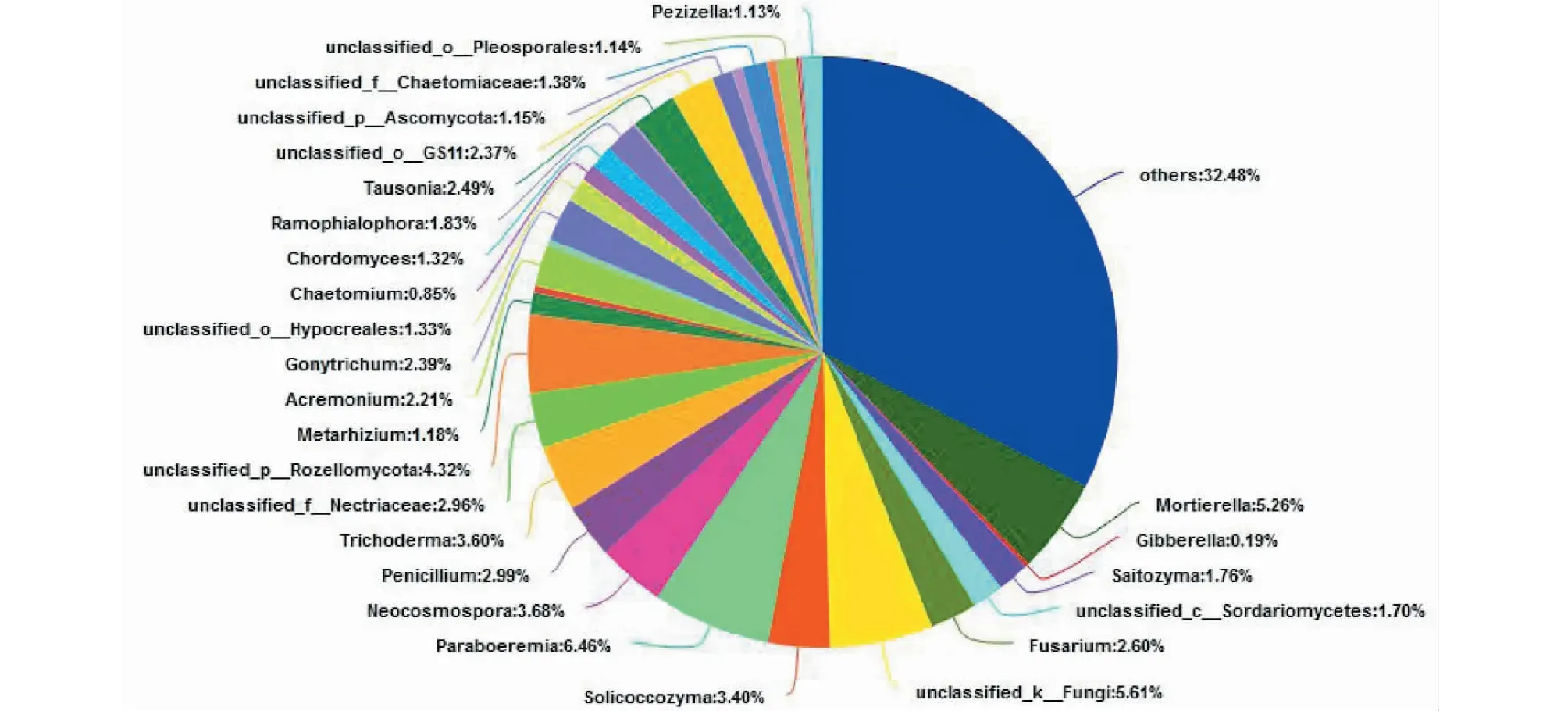
图9 C样品属水平真菌群落组成
2.3 组间差异比较分析
由样品菌落构成分析和物种Venn图(图10)可知,3组采集样品真菌群落多样性较高,真菌群落结构差异性适中,存在共有和特有物种。3组样品真菌物种广泛存在担子菌门(Basidiomycota)、子囊菌门(Ascomycota)、被孢霉菌门(Mortierellomycota)3个门类。A样品OTU数量为1704个,B样品OTU数量为1204个,C样品OTU数量为2118个。A组样品与B组样品共有384个OTUs,A组样品与C组样品共有640个OTUs,B组样品与C组样品共有515个OTUs,3组样品共有283个OTUs。A组样品和B组样品共有优势真菌属:赤霉属(Gibberella)、镰刀霉属(Fusarium)、被孢霉属(Mortierella)、毛壳菌属(Chaetomium)。B组样品和C组样品共有优势真菌属:镰刀霉属(Fusarium)、支顶孢属(Acremonium)、新赤壳属(Neocosmospora)、被孢霉属(Mortierella)、Ramophialophora属、Chordomyces属。A组样品和C组样品共有优势真菌属:被孢霉属(Mortierella)、木霉属(Trichoderma)、青霉属(Penicillium)、镰刀霉属(Fusarium)、Solicoccozyma属、Saitozyma属。三者共有优势真菌属为被孢霉属(Mortierella)和镰刀霉属(Fusarium)。
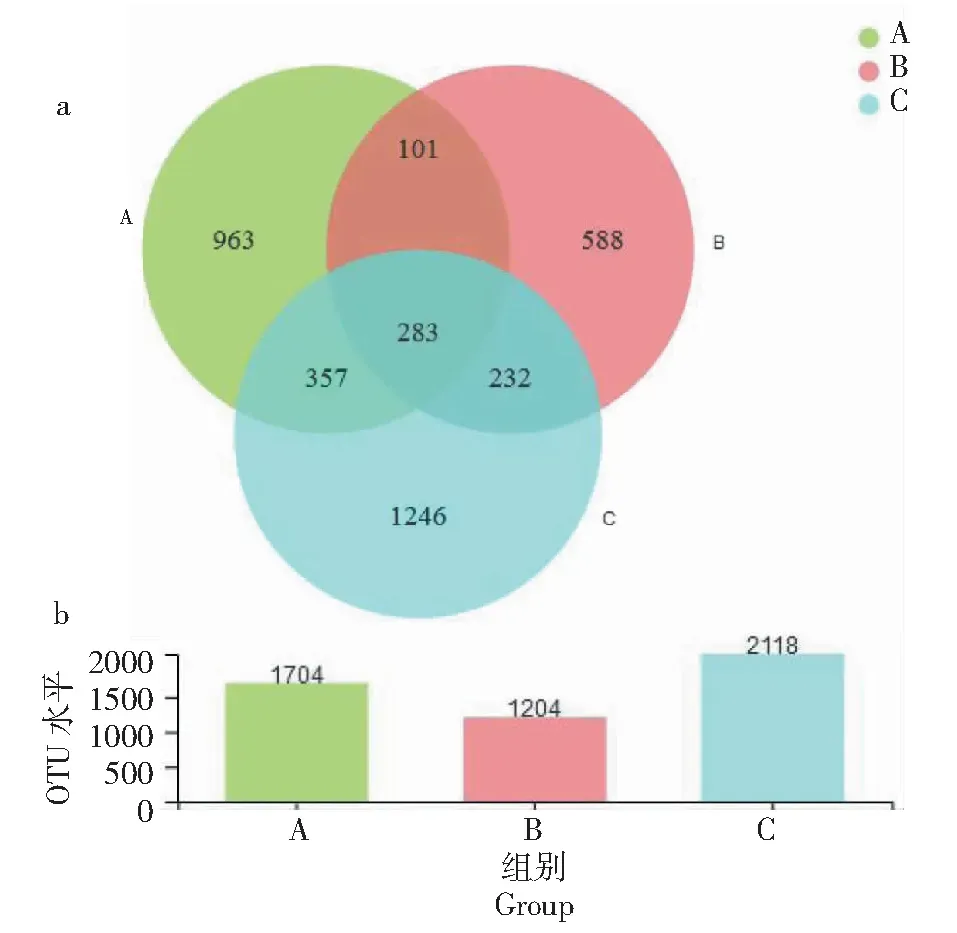
图10 不同地区样品Venn分析
基于3组样品中群落丰度数据,运用严格的统计学方法检测不同样品真菌群落中表现出的丰度差异的物种,进行假设性检验,评估观察到的差异的显著性。对比3组样品真菌群落丰度水平,结果显示被孢霉属、赤霉属、Saitozyma属、Solicoccozyma属、Paraboeremia属、青霉属、木霉属等菌属在3组样品中群落丰度差异较大,其中赤霉属、木霉属真菌群落丰度在3组样品中存在显著性差异(P<0.05,图11)。
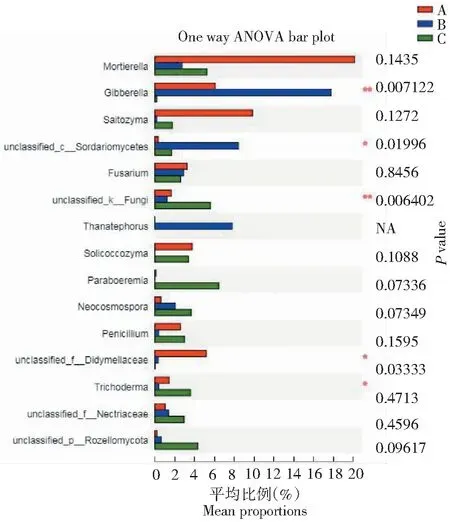
*表示差异显著,**表示差异极显著
2.4 真菌群落功能预测分析
如表6所示,通过KEGG数据库比对分析,真菌参与了多种代谢功能,其中包含β-甘露聚糖酶、α-葡聚糖合酶、α-甘露聚糖酶等功能,为以后筛选或指导微生物利用提供帮助。
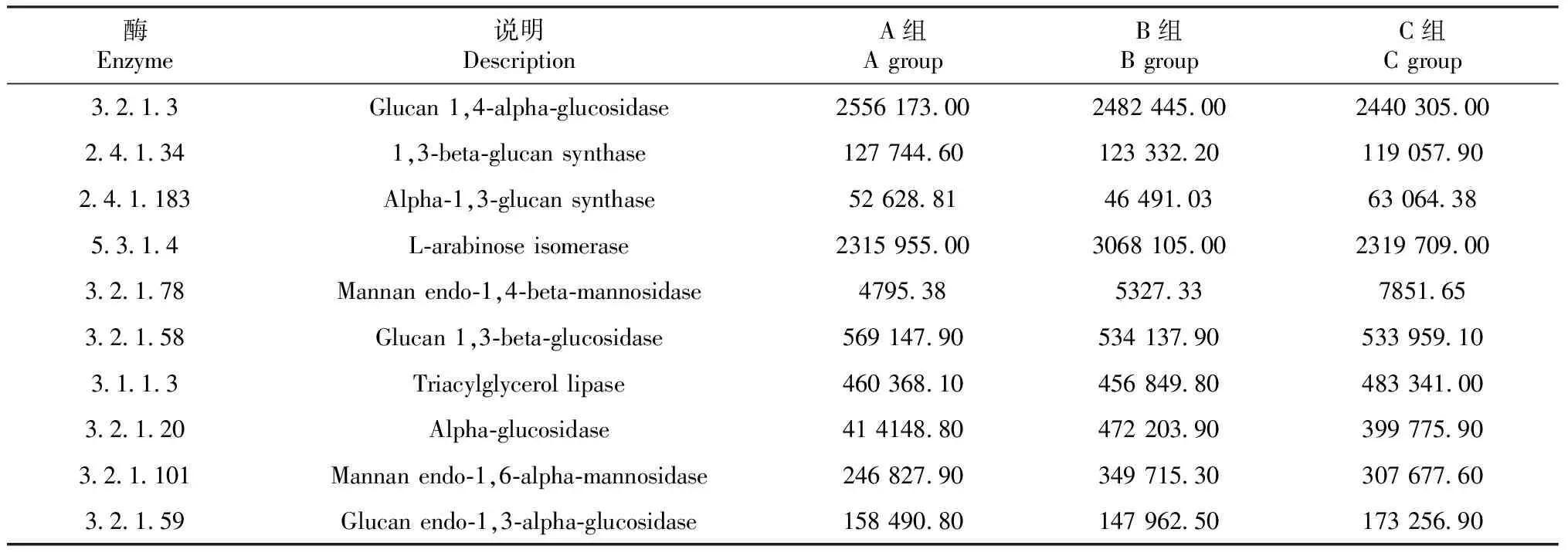
表6 不同地区样品真菌群落KEGG功能丰度统计
3 讨 论
通过利用高通量测序技术对三地区样品真菌群落多样性进行了分析,共得到3770个OTUs,获得真菌菌落17门、62纲、141目、326科、747属、1302种。C样品的香农指数较高,辛普森指数较低,Chao1 指数和ACE指数高,表明C样品真菌群落多样性和丰富度较高。物种组成分析结果表明不同地区魔芋根际土壤真菌群落存在差异,赤霉属、木霉属真菌群落丰度在3组样品中存在显著差异(P<0.05)。3组采集样品真菌群落多样性较高,A组样品与B组样品共有384个OTUs,A组样品与C组样品共有640个OTUs,B组样品与C组样品共有515个OTUs,3组样品共有283个OTUs。C样品真菌参与了多种代谢功能,其中包含β-甘露聚糖酶、α-葡聚糖合酶、α-甘露聚糖酶等功能,为以后筛选或者定向指导微生物利用提供帮助。3组样品对比分析发现,种植年限对魔芋种魔芋根际土壤真菌群落多样性影响较大,而海拔高度和经纬度对其影响较小。
4 结 论
通过利用高通量测序技术对三地区真菌群落多样性进行了分析,对比分析结果发现不同地区魔芋根际土壤真菌群落差异较大,种植年限对魔芋根际土壤真菌群落多样性水平影响较大,而海拔高度和经纬度对魔芋根际土壤真菌群落多样性水平影响较小,为魔芋的产业发展提供了帮助。
猜你喜欢
当代水产(2022年8期)2022-09-20
中国音乐学(2022年2期)2022-08-10
农业工程学报(2022年8期)2022-08-08
中国土壤与肥料(2022年4期)2022-06-14
草地学报(2022年4期)2022-04-25
昆明医科大学学报(2022年2期)2022-03-29
土壤学报(2022年1期)2022-03-08
学校教育研究(2020年7期)2020-04-09
家庭医药(2018年5期)2018-05-22
家庭百事通·健康一点通(2018年3期)2018-03-30
