
CD24调节大肠癌中Wnt/β-catenin信号转导
2022-11-17 12:33邓启亮郑欢欢
健康之友 2022年21期
邓启亮 郑欢欢
(1 吉安市中心人民医院/消化内科 江西 吉安 343000 2 吉安市中心人民医院/内分泌科 江西 吉安 343000)
CD24是一种与膜连接[1]的糖基磷脂酰肌醇(GPI)锚定蛋白。CD24可以影响整合素β1的侧向定位,并将整合素β1募集到脂筏中。Wnt/β-catenin是细胞-细胞粘附的胞质组分,它与E-cadherin胞质结构域相互作用,并通过α-catenin与肌动蛋白细胞骨架连接。众所周知,β-catenin是Wnt通路中的一个关键分子,当Wnt通路被激活时,β-catenin在细胞质中积累,然后转运到细胞核。β-catenin在Tcf/LEF位点与DNA结合,并通过与Tcf/LEF位点结合诱导下游基因靶点如cyclin D1[2]和c-myc[3]的转录和表达。在以前的研究确定了CD24在结肠癌细胞粘附、增殖及转移中的作用,但尚未阐明其潜在的机制。
1 材料和方法
1.1抗体和化学药品:使用以下抗体:抗cyclinD1、c-myc、GSK-3β、P-GSK-3β的抗体购自CST(细胞信号技术公司,Beverly,MA)。抗β-catenin和抗β-actin购自Santa Cruz Biotechnology(Santa Cruz Biotechnology,CA)。TCF/LEF报告者TOP flash及其突变体控制物FOP flash是从美国北部(纽约市)购买的
1.2免疫印迹分析:培养和收集的细胞在添加蛋白酶抑制剂的RIPA(Beyotime,shanghai,China)缓冲液中裂解。裂解物在4°C下离心30分钟。为了确定β-catenin在细胞中的定位,用细胞分馏试剂盒(Keygen,南京,中国)从细胞中分离细胞质和核蛋白。等量蛋白质样品在分离凝胶中用SDS-PAGE电泳分离,转移到硝化纤维素片上。在5%脱脂牛奶中阻断2小时后与稀释后的原抗体(cyclinD1 1:1000,c-myc 1:500,GSK-3β1:1000,P-GSK-3β1:1000,β-catenin 1:200,β-actin 1:1000)在4℃下孵育过夜。用TBS-T洗膜,并与辣根过氧化物酶结合的二级抗体(1:5000稀释度)一起孵育,使用ECL检测试剂盒(Pierce Biotechnology Inc.,Rockford,IL)加强化学发光检测。
1.3免疫荧光素:免疫荧光素使用以下一抗:小鼠抗β-catenin(1:400)。二次抗体为山羊抗小鼠FITC(1:1000)。染色切片用荧光显微镜观察和拍照。
1.4瞬时转染和荧光素酶试验:瞬时转染试验将细胞一式三份接种在24孔板中。用Lipofectamine 2000共转染萤火虫荧光素酶报告构建体TOP flash或FOP flash和含有肾母荧光素酶的参考构建体pRL-SV40(Promega,Madison,WI)。48小时后,在GloMax发光测量系统(Promega,Madison,WI)中使用双荧光素酶报告分析系统试剂盒(Promega,Madison,WI)测量荧光素酶活性。以荧光素酶报告基因转染SW480-CD24和SW480-V细胞或SW620-KD和SW620-NC,共转染pRL-SV40报告基因作为内对照,将萤火虫荧光素酶活性与肾母细胞荧光素酶活性进行归一化,并以平均±SD作图。
2 结果
2.1 CD24的表达改变β-catenin表达量并改变其定位
从SW480-CD24和SW480-V细胞中制备细胞质蛋白和核蛋白,并对其进行免疫印迹分析,以确定CD24是否调节β-catenin蛋白的表达。与SW480-V细胞相比,SW480-CD24克隆的胞浆β-catenin水平增加了40%以上,细胞核中的β-catenin水平增加了80%以上(图.1C)。由于β-catenin在信号转导中的功能依赖于其胞质/核定位,进一步通过间接免疫荧光分析了β-catenin蛋白的分布。β-catenin在细胞质中染色增强(图.1A)。结果表明,CD24的上调导致SW480细胞β-catenin表达量增加并使其从细胞膜向细胞质转移。
siRNA敲除CD24降低β-catenin表达并改变其定位,与过度表达CD24的SW480细胞相比,SW620-KD的细胞质β-catenin水平比SW620-NC细胞下降了80%,细胞核下降了40%(图.1D)。进一步通过间接免疫荧光分析了β-catenin蛋白的分布。β-catenin在细胞质中染色减弱(图1.B)。结果表明,在SW620细胞中,CD24下调导致β-catenin表达量减少并使其从细胞质向细胞膜转移。

图1 CD24的表达改变β-catenin表达量并改变其定位
2.2 CD24可调控β-catenin/TCF/LEF信号转录活性
CD24过表达可激活β-catenin/TCF/LEF信号转导通路。CD24表达上调增加了SW480细胞质和细胞核中β-catenin的表达,并导致β-catenin从细胞膜向细胞质的重新定位。使用TOP flash和FOP flash检测了SW480-CD24和SW480-V的β-catenin/TCF/LEF信号。如(图2.A)所示,SW480-CD24细胞的β-catenin/TCF/LEF复合物的相对转录活性比SW480-V细胞提高了4~6倍。
沉默CD24可抑制β-catenin/TCF/LEF信号转导通路。SW620-KD细胞中β-catenin/TCF/LEF信号通路如(图2.B)所示,SW620-KD细胞的β-catenin/TCF/LEF复合物的相对转录活性比SW620-NC细胞降低了3~4倍。
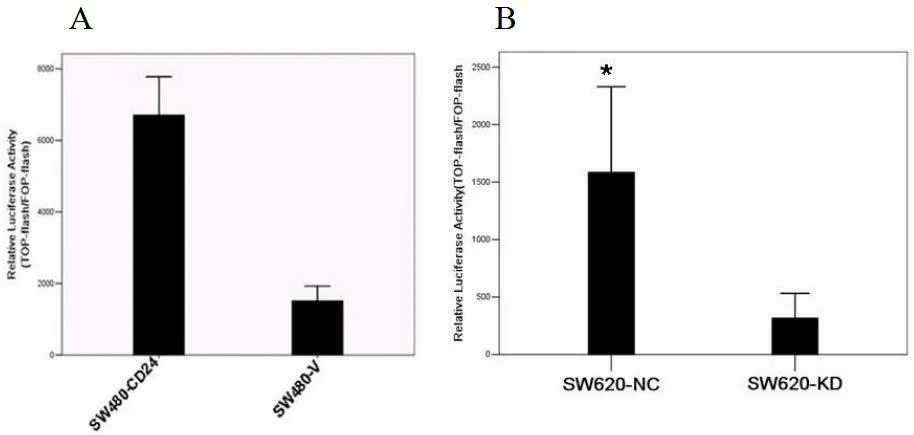
图2 CD24可调控β-catenin/TCF/LEF信号转录活性
2.3 CD24通过GSK3-β磷酸化调节cyclinD1、c-myc的表达。
为了解CD24是否影响TCF/LEF的下游信号通路,随后检测了GSK3-β和GSK3-β的磷酸化水平,数据表明(图3.A)过表达CD24可以诱导GSK3-β的磷酸化。与β-catenin/TCF/LEF活性表达一致,在SW480-CD24中c-myc和cyclinD1的表达量随CD24的增加而升高,而在SW620-KD中GSK3-β的磷酸化水平下降,c-myc和cyclinD1的表达量随之减少(图3.B)。

图3 CD24通过GSK3-β磷酸化调节cyclinD1、c-myc的表达
3 讨论
以前的研究表明,在MTLyCD24mut中CD24的重新表达可以通过激活整合素促进与细胞外基质成分的结合[4]。β-catenin表达和信号转导异常是多种恶性肿瘤尤其是结直肠癌发生的重要因素。研究中发现SW480-CD24细胞胞质和核β-catenin蛋白水平明显升高,CD24的过度表达使β-catenin从细胞膜向胞质和细胞核易位导致β-catenin/TCF/LEF信号转导增加。而沉默CD24可使β-catenin从细胞质和细胞核向细胞膜易位导致β-catenin/TCF/LEF信号转导减弱。
我们进一步研究了β-catenin/TCF/LEF转录活性所反映的Wnt信号转导是否与β-catenin表达增加和定位改变有关。Tcf/LEF荧光素酶报告构建是检测β-catenin/Tcf/LEF信号转导转录激活的有效方法[5],通过计算结肠癌细胞中TOPflash与FOPflash荧光素酶活性的比值,证实了结肠癌细胞中β-catenin/Tcf/LEF转录活性与CD24的表达相关。
本研究所研究的Wnt途径的另一个重要成分是GSK-3β,通常胞质β-catenin库是由GSK-3β和其他破坏复合物的成分维持的,GSK-3β的磷酸化可以破坏泛素-蛋白体系统,导致胞质β-catenin库和核库的增加[6,7],我们发现CD24可以诱导GSK3-β的磷酸化。以往研究还发现CD24能激活intergrinβ1,整合素[8]能激活多种蛋白酪氨酸激酶,包括粘着斑激酶(FAK)、Src家族激酶,而GTPase Cdc42 GTPase Cdc42通过激活PI3K/AKT使GSK3-β磷酸化,从而抑制GSK3-β的表达,推测intergrinβ1与GSK3-β可能通过GTPase Cdc42存在一定的关系。
总之,我们的数据表明CD24可调控Wnt/β-catenin转录活性,使β-catenin表达量改变并在细胞膜、细胞质及细胞核中易位。CD24为结肠癌的治疗提供了可能的策略。
猜你喜欢
临床与实验病理学杂志(2022年10期)2022-12-19
蔬菜(2022年8期)2022-08-25
课外生活·趣知识(2022年2期)2022-02-08
贵州农业科学(2021年7期)2021-09-13
湖南农业大学学报(自然科学版)(2021年4期)2021-08-14
云南医药(2021年3期)2021-07-21
高考·中(2019年6期)2019-09-10
江苏农业科学(2017年14期)2017-10-10
吉林农业(2017年5期)2017-05-13
科学中国人(2016年30期)2016-07-14
