
谷子TGA转录因子家族全基因组鉴定与分析
2023-05-15 01:27秦迎迎杨致荣
山西农业科学 2023年5期
高 倩 ,秦迎迎 ,杨致荣
(1.山西农业大学 基础部,山西 太谷 030801;2.山西农业大学 生命科学学院,山西 太谷 030801)
谷子(Setaria italica)起源于我国,是我国重要 的杂粮作物之一,在漫长的中国北方旱地农耕文明历史中占有重要地位。谷子属于1年生草本植物,是典型的二倍体禾本科作物(2n=18),因其抗旱、耐贫瘠、适应性广等特性,在当前的种植业结构调整中起着重要作用。谷子脱壳后称为小米,是重要的健康保健食品,不仅营养价值丰富,而且可以改善慢性消化系统疾病[1-3]。最新研究也发现,小米群体代谢组与抗炎活性之间具有一定的关联性,为小米作为药食同源的食材和服务大健康产业提供了理论支撑[4]。
近年来,植物中越来越多的转录因子(Transcription factor,TF)被发现与研究。转录因子能够与靶基因启动子上的顺式作用元件相互作用,激活或抑制下游靶基因表达,在植物生长发育以及胁迫响应过程中具有关键功能[2-3,5]。碱性亮氨酸拉链(Basic region/leucine zipper,bZIP)转录因子参与植物的生长发育、病原防御、高温以及干旱胁迫应答等生物和非生物胁迫反应,根据其结构和功能被分为A、B、C、D、E、F、G、H、I和S共10个亚族[1,6]。TGA(TGACG motif-binding factor)转录因子属于bZIP转录因子家族中的D亚族[6],含有碱性区域(Basic region)和亮氨酸拉链(Leucine-zipper region)功能域[7],能够特异性地与TGACG激活序列(Activation sequence 1,as-1)结合来调控下游靶基因的表达,参与防御病害和生理生长2种不同进程[7-9]。TGA首先在烟草中被发现和克隆[10],之后在 拟 南 芥[5,11]、玉 米[9]、水 稻[12]、大 豆[13]、大 麦[14]等 植物中被发现和研究。在拟南芥中,TGA家族基因有10个成员,分为Ⅰ~Ⅴ共5个亚组[5,11],含有1~3个基因,其中亚组Ⅰ、亚组Ⅱ和亚组Ⅲ成员参与拟南芥响应水杨酸(Salicylic acid,SA)调控植株的抗病反应,亚组I、亚组Ⅱ和亚组Ⅴ成员参与花器官的发育过程,TGA8/PAN在拟南芥根尖静止中心细胞中表达,与亚组Ⅱ的成员共同调控根的生长发育,在干细胞群动态平衡、细胞氧化还原动态平衡、植株抵抗病菌和光合作用中共同发挥作用[5]。在玉米叶的生长过程中,bZIP家族D亚族的Liguleless2基因能够定位玉米叶片和叶鞘的边界[9]。柯丹霞等[13]研究发现,在大豆中,GmTGA26可以调控下游的一系列胁迫响应基因,在很大程度上增强植株的耐盐能力。连梓伊等[12]和李欢鹏等[14]研究发现,水稻和大麦中TGA转录因子与拟南芥类似,能够响应SA调控的抗病反应,从而提高植株抗病性[15-18]。然而,目前在谷子中有关TGA家族基因成员及其功能的研究未见报道。
谷子具有基因组小(430 Mb)、自花授粉、生长周期短等特点,已经逐渐成为C4禾谷类模式植物[19]。本研究通过以谷子超早熟突变体xiaomi为研究对象,对谷子的TGA家族基因(SiTGAs)进行全基因组鉴定和初步预测分析,旨在为进一步研究谷子TGA转录因子的生物学功能提供理论依据。
1 材料和方法
1.1 谷子TGA家族基因序列鉴定及其理化性质分析
在拟南芥数据库TAIR(https://www.arabidopsis.org/)中搜索获得拟南芥TGA的氨基酸序列,之后在谷子xiaomi数据库MDSi:Multi-omics Database forSetaria italica(http://foxtail-millet.biocloud.net/home)中利用BlastP筛选xiaomi的TGA基因,筛选参数为score value≥100,e-value≤1×10-10,并利用Pfam(http://pfam.xfam.org/)对得到的氨基酸序列进行结构域验证。最后利用EXPASY-ProtParam在线工具(https://web.expasy.org/protparam/)预测谷子TGA基因家族成员蛋白质一级结构的理化性质,包括氨基酸的数目、相对分子质量、等电点等,使用PSORT Prediction(http://psort1.hgc.jp/form.html)在线网站进行亚细胞定位,预测TGA家族蛋白质的表达位置。
1.2 谷子TGA家族基因的染色体定位
根据MDSi数据库中xiaomi基因组文件(.gff格式)中的注释信息,使用TBtools软件提取谷子TGA家族基因在染色体上的位置信息,并对其进行可视化分析。
1.3 谷子TGA家族基因蛋白质的二级结构
将谷子TGA家族基因蛋白的氨基酸序列提交至SOPMA在线网站(http://www.prabi.fr/),分析其蛋白质二级结构。
1.4 谷子TGA家族基因的系统进化树构建
使用Phytozome在线网站(https://phytozome.jgi.doe.gov/pz/portal.html)和甘蔗基因组数据库(http://sugarcane.zhangjisenlab.cn/sgd/html/downl oad.html),以TGA为关键词,搜索并下载水稻、高粱和甘蔗TGA家族成员的氨基酸序列。使用MEGA 7.0软件中的Align by ClustalW程序对谷子和拟南芥、水稻、高粱、甘蔗TGA基因家族成员的氨基酸序列进行多重比对,并采用邻接法(Neighbor-Joining,NJ)构建系统进化树,Bootstrap参数设置为1000。
1.5 谷子TGA家族的基因结构分析
分析并整理xiaomi基因组信息(.gff格式),在Gene Structure Display Server(GSDS,http://gsds.cbi.pku.edu.cn/)在线软件中选择.gff模式进行TGA家族基因结构图的绘制。
1.6 谷子TGA家族基因保守基序及保守结构域分析
将谷子TGA家族基因成员的蛋白序列提交至MEME在线软件(https://meme-suite.org/meme/tools/meme),预测其保守基序,输出MAST MXL文件后,使用TBtools的MEME/MAST motif pattern redrawer功能进行可视化处理,绘制TGA家族基因Motif示意图。使用NCBI-CDD数据库(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)的Batch-CD-Search功能进行批量查询,确定谷子TGA家族基因成员氨基酸序列的保守结构域,再利用TBtools软件进行可视化分析。
1.7 谷子TGA家族基因启动子分析
利用MDSi数据库获得谷子TGA家族基因的启动子区域(ATG上游的2000 bp序列),将其提交至在线数据库PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),分析并确定谷子TGA家族基因启动子区顺式作用元件,使用TBtools软件对其进行可视化分析。
1.8 谷子TGA家族基因表达分析
利用MDSi数据库中获取谷子TGA家族基因不同时期不同组织中的表达数据,将其提交至Heatmapper(http://www.heatmapper.ca/expression/)在线软件中,分析并绘制TGA家族基因特异的组织表达热图。
2 结果与分析
2.1 谷子TGA家族基因成员的鉴定及其理化性质分析
通过同源比对,在谷子xiaomi中共鉴定到16个TGA家族基因成员,根据其在染色体上的排布,命名为SiTGA1~SiTGA16(图1),对应的基因名称及其理化性质如表1所示。SiTGAs在谷子的9条染色体上均有分布,SiTGA1、SiTGA14、SiTGA15、SiTGA16分别分布于1、7、8号和9号染色体上;SiTGA2-4、SiTGA9-11分别分布于2号和5号染色体上;SiTGA5-6、SiTGA7-8、SiTGA12-13则分别分布于3、4号和6号染色体上(图1)。
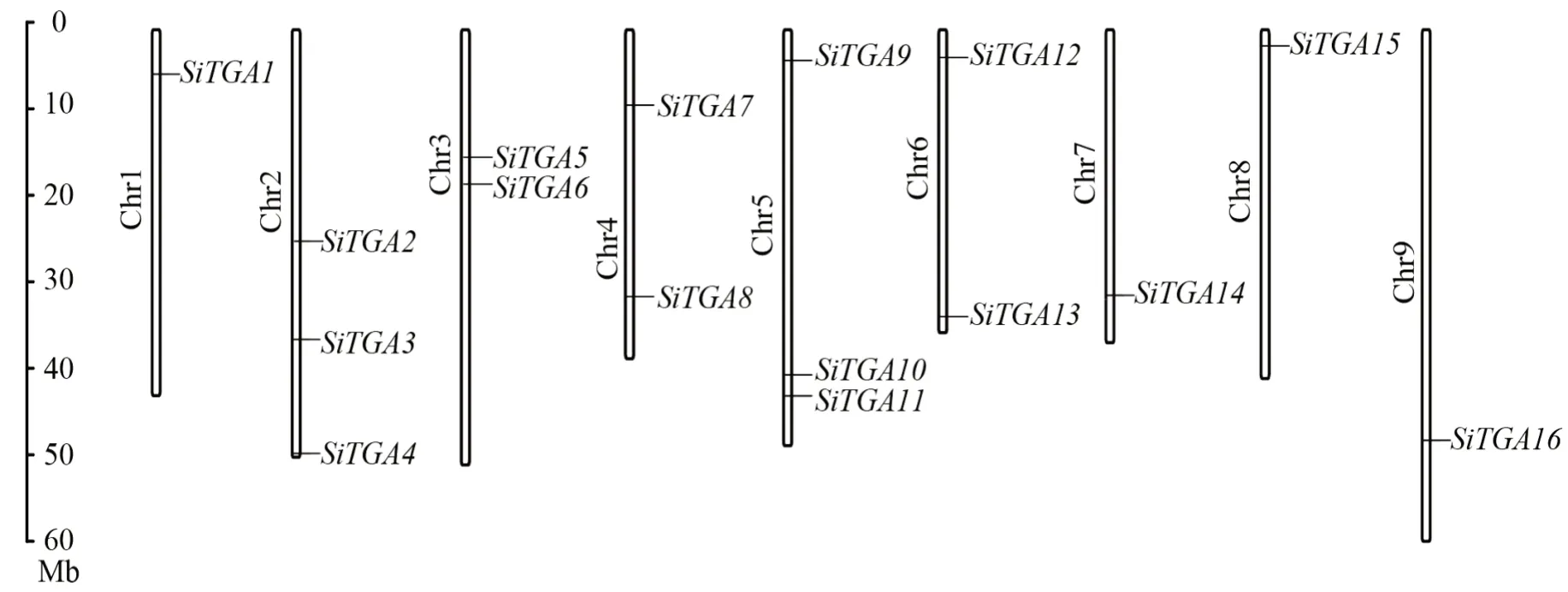
图1 谷子TGA基因家族成员染色体定位Fig.1 Chromosome localization of TGA gene family members in Setaria italica
对谷子TGA家族基因理化性质进行分析可知(表1),谷子TGA家族基因成员的基因序列长度为1906(SiTGA5)~8823 bp(SiTGA2),CDS序列长度996~1617 bp;蛋白质分子质量为36562.32~58030.21 u;蛋白质等电点范围为6.18(SiTGA10)~9.25(SiTGA8),其中碱性蛋白8个,酸性蛋白8个。
从表2可以看出,谷子TGA基因家族蛋白二级结构中,α-螺旋为主要结构,占比为49.59%~68.88%;无规卷曲次之,占比为22.39%~41.98%;β-折叠和直链延伸占比较少,仅为1.45%~6.15%和4.45%~9.94%。对谷子TGA家族基因成员进行亚细胞定位预测可知(表1),SiTGA9定位在质膜表达,SiTGA13定位在内质网膜表达,其他的14个谷子TGA家族基因成员都定位于细胞核内,这与其主要作为转录因子发挥作用一致。

表1 谷子TGA基因家族的基本信息Tab.1 Basic information of TGA gene family in Setaria italica

表2 谷子TGA基因家族成员二级结构占比Tab.2 Proportion of secondary structure of TGA gene family members in Setaria italica %
2.2 谷子TGA家族基因成员系统进化及基因结构分析
为了研究谷子TGA家族基因的进化关系,对拟南芥的10个AtTGAs、水稻的15个OsbZIPs、高粱的22个TGA家族基因成员(Sobic.001G243800~Sobic.010G192432)、甘蔗的3个TGA家族基因成员(Sof008184、Sof001897、Sof009733)和 谷 子 的16个SiTGAs进行系统进化树的构建,结果如图2所示,谷子SiTGAs家族种间进化树可分为6个亚组,分别是亚组Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ,其中SiTGA1、SiTGA2、SiTGA3、SiTGA8属 于 第Ⅰ亚 组,与AtTGA10聚为一支,与第Ⅱ亚组的SiTGA6、SiTGA11、SiTGA13、SiTGA15聚为一支;SiTGA4、SiTGA5、SiTGA9、SiTGA10、SiTGA16隶属于第Ⅳ亚组,其中SiTGA10与OsbZIP08聚为一支;亚组Ⅳ与亚组Ⅲ的SiTGA7聚为一支;亚组Ⅴ的SiTGA14和亚组Ⅵ的SiTGA12聚为一支。整体来看,谷子与高粱的亲缘关系最近。

图2 拟南芥、水稻、高粱、甘蔗和谷子中TGA基因家族的系统进化分析Fig.2 Phylogenetic analysis of TGA gene family in Arabidopsis thaliana,Oryza sativa,Sorghum bicolor,Saccharum officinarum,and Setaria italica
对谷子TGA家族基因成员的基因结构进行分析,结果显示(图3),全部基因呈现断裂基因特征,均包含有外显子和内含子;16个SiTGAs基因结构存 在 一 定 的 差 异,SiTGA1、SiTGA5、SiTGA9、SiTGA12、SiTGA13、SiTGA15基因结构中不含有5′和3′端非编码区(UTR),其余的SiTGAs均含有非 编 码 区,其 中SiTGA4、SiTGA6、SiTGA11、SiTGA14、SiTGA16含有2~3个5′端非编码区(UTR),SiTGA8含有2个3′端非编码区(UTR)。各基因的外显子数目有7~12个不等,SiTGA11、SiTGA13、SiTGA15的外显子数目较多,分别有12个,而SiTGA12的外显子仅有7个。

图3 谷子TGA家族基因成员基因结构Fig.3 Gene structure map of TGA gene family members in Setaria italica
2.3 谷子TGA家族基因成员保守基序及保守结构域分析
为了进一步了解谷子TGA家族基因成员的可能发挥的功能,对其保守基序和保守结构域进行了分析,结果发现(图4),除SiTGA2不含有Motif 7基序,其余SiTGAs都含有Motif 1~Motif 7。Motif 8基序为亚组Ⅰ和亚族Ⅱ所特有;Motif 9和Motif 10基序为亚组Ⅳ所特有,说明Motif 8~Motif 10这3种基序在进化中分化程度很低,这可能是SiTGAs不同亚组在不同生理过程中发挥作用的重要原因。

图4 谷子TGA家族基因成员保守基序(左)和保守结构域(右)分析Fig.4 Analysis of conserved motifs(left)and conserved domains(right)of TGA gene family members in Setaria italica
谷子TGA家族基因成员共包含4个保守结构域:DOG1、bZIP-HBP1b-like、bZIP superfamily和PHA03418 superfamily,每个SiTGAs都含有2~3个保守结构域。进一步分析发现,16个SiTGAs都含有DOG1结构域,DOG1结构域能够高度特异性地控制种子休眠,推测谷子TGA家族基因成员能够特异性地调控种子休眠。只有SiTGA1含有PHA03418 superfamily结构域,推测SiTGA1有不同于其余SiTGAs的功能。SiTGA7、SiTGA10、SiTGA13、SiTGA14、SiTGA15都含有bZIP_HBP 1b-like和DOG1保守结构域。SiTGA4、SiTGA5、SiTGA9、SiTGA16所含保守结构域的位置相似,推测它们具有相似的生物学功能。亚族Ⅰ成员除SiTGA1外其余3个都含有相同的保守结构域,亚族Ⅳ除SiTGA10外保守结构域的种类也相同,说明谷子TGA家族基因不同亚族间成员的保守结构域具有保守性。
2.4 谷子TGA家族基因成员启动子序列分析
为了更深入地了解谷子SiTGAs的潜在功能,利用PlantCARE网站对SiTGAs的启动子顺式作用元件进行分析,共鉴定出17类顺式作用元件,结果如图5所示。
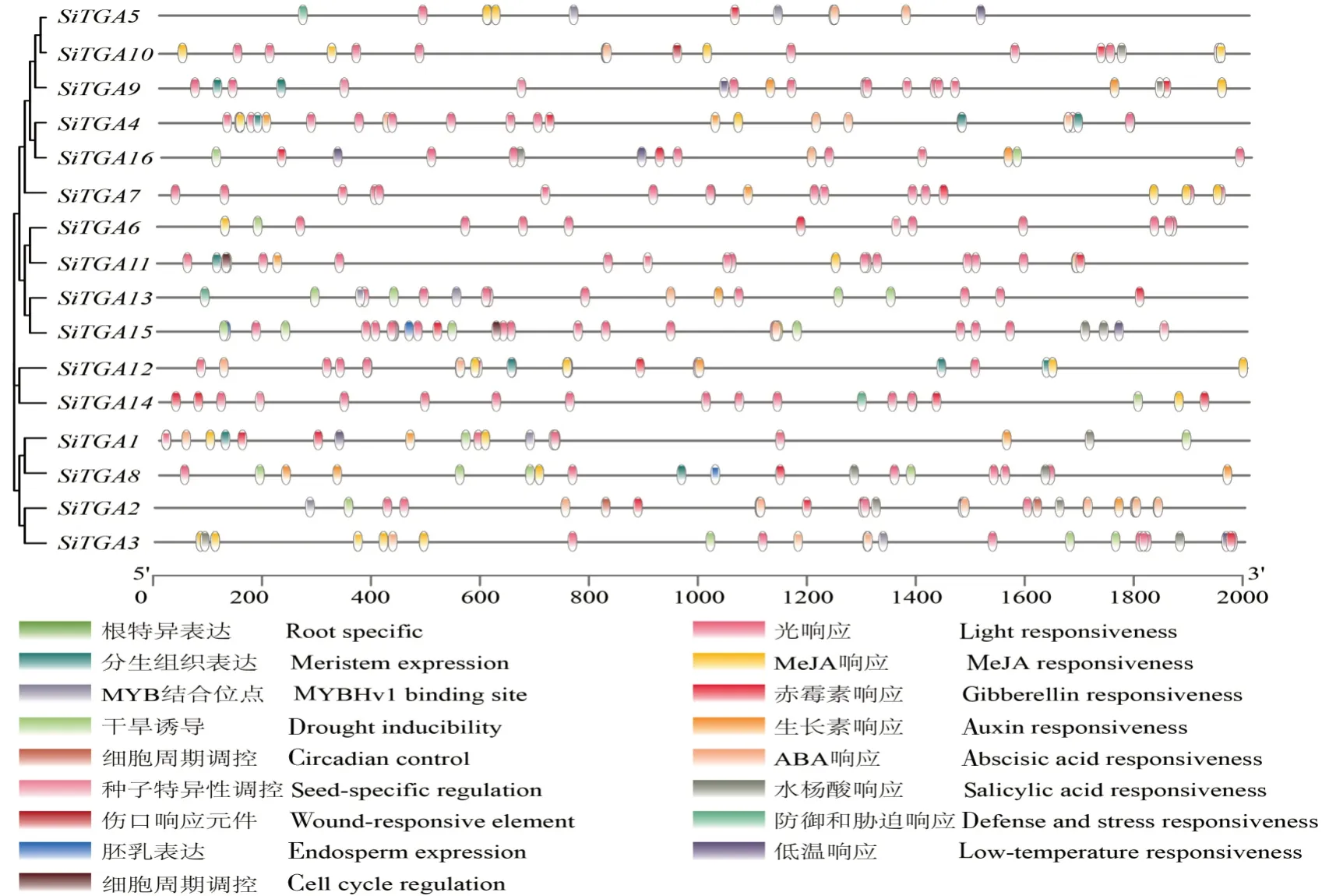
图5 谷子TGA家族基因成员启动子顺式作用元件Fig.5 Cis-acting elements of TGA gene family member promoter in Setaria italica
从图5可以看出,谷子TGA家族基因各成员具有顺式作用元件类型的数量为4(SiTGA7)~11(SiTGA1)个。所有的谷子TGA家族基因成员都有光反应元件和赤霉素反应元件,推测SiTGAs基因的表达受到光诱导的调节,而且在赤霉素响应中也发挥着作用。大多数SiTGAs启动子中含有水杨酸响应元件、ABA响应元件、干旱诱导元件,说明SiTGAs参与谷子的抗病性和抗逆性调控。除此之外,根特异的响应元件只存在于SiTGA1启动子中,推测SiTGA1参与根分化过程的调控;生物钟调控元件只存在于SiTGA2中,推测该基因可能参与昼夜节律的调控;SiTGA11、SiTGA13有细胞周期调控元件,这2个基因可能参与细胞周期的调控。
2.5 谷子TGA家族基因成员表达分析
为了探究SiTGAs在谷子生长发育过程中是如何发挥功能的,根据其在谷子不同组织以及发育不同时期的表达模式,绘制了基因表达热图如图6所示,SiTGA7在花序中有较高的表达量;SiTGA3和SiTGA5表达模式比较类似,在萌发3 d的籽粒、未成熟小穗期和未成熟籽粒期表达量较高;除SiTGA3、SiTGA5、SiTGA7以外,其余的13个基因都在灌浆期根中有较高的表达。除此之外,SiTGA10在灌浆期穗颈节间和成熟后的籽粒中表达量较高,SiTGA15在萌发3 d的籽粒中表达量较高,SiTGA9、SiTGA2、SiTGA14、SiTGA1、SiTGA8、SiTGA4、SiTGA16在谷子灌浆期表达量较高。
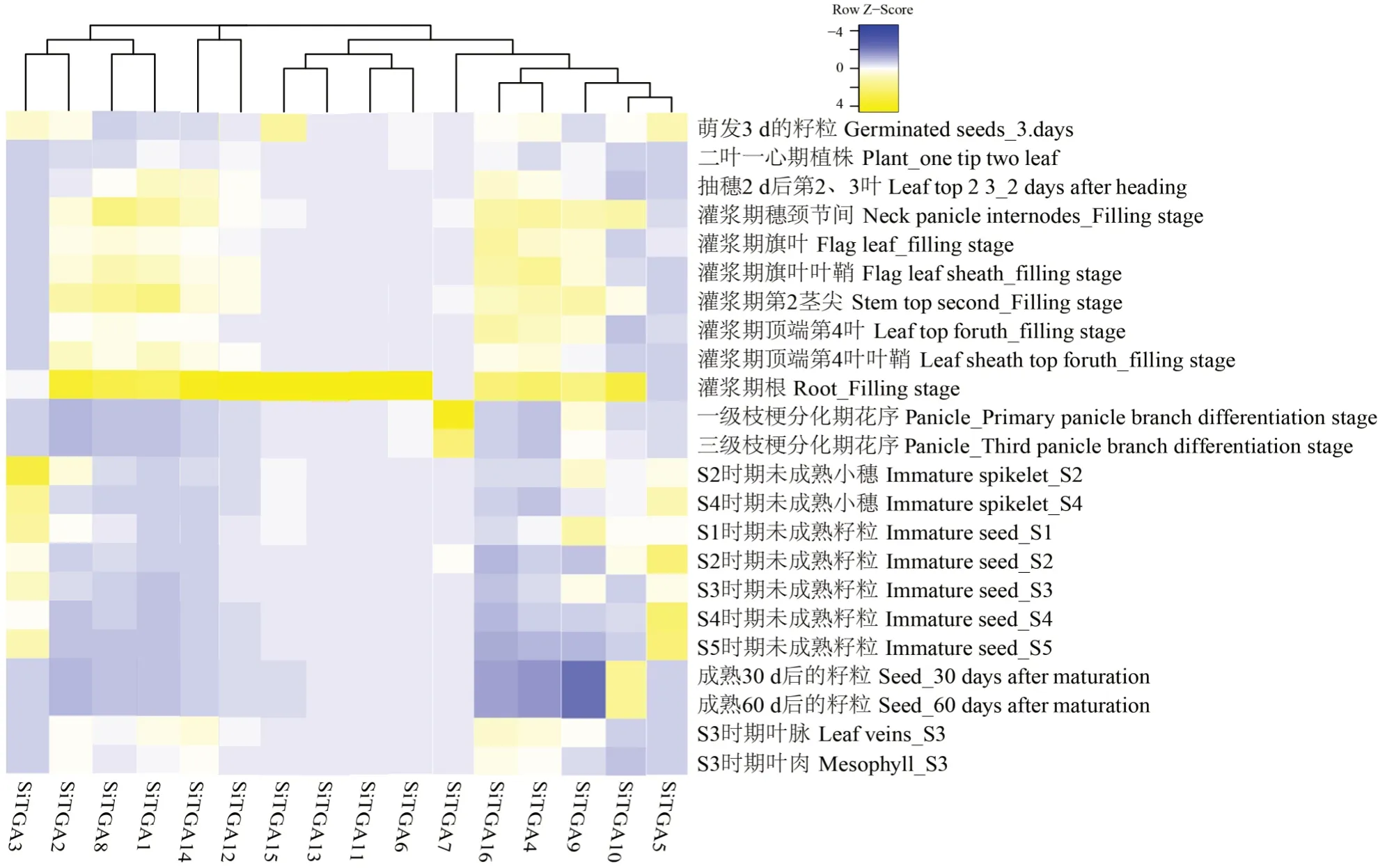
图6 谷子TGA家族基因组织表达模式Fig.6 Tissue expression pattern of TGA gene family in Setaria italica
3 结论与讨论
本研究以谷子超早熟材料xiaomi为研究对象,通过与拟南芥中的TGA家族基因的同源比对,筛选得到了谷子的16个SiTGAs。通过系统进化分析发现,SiTGA1、SiTGA2、SiTGA3、SiTGA8和拟南芥的AtTGA10聚在Ⅰ亚组,这4个SiTGAs的保守基序和保守结构域总体较为相似,但是也有一些差异,其中SiTGA2中缺少Motif 7,SiTGA1缺少非编码区且特有PHA03418 superfamily保守结构域。AtTGA10具有调控植株抗病和花器官发育的双重作用[20-21],而组织表达结果显示,只有SiTGA3在未成熟的小穗和籽粒中表达量较高,其余3个SiTGAs在灌浆期尤其是灌浆期根中表达量较高,说明其功能与拟南芥中不完全相同,在谷子根的生长发育过程中发挥重要作用。
亚组Ⅱ成员SiTGA6、SiTGA11、SiTGA13和SiTGA15的保守基序种类相同,但SiTGA13、SiTGA15不含有非编码区;SiTGA6和SiTGA11含有bZIP superfamily保守结构域,而SiTGA13和SiTGA15含有bZIP_HBP1b-like结构域。该亚组成员主要在灌浆期根部表达,推测其在谷子根发育中发挥重要作用。SiTGA15在萌发3 d的籽粒中表达也较高,说明其也参与调控籽粒萌发过程。同源基因OsbZIP79可影响萜类植保素积累从而调控水稻的抗病性[22],推测该亚组成员参与谷子的抗病性。
SiTGA7和AtTGA8以及OsbZIP47聚为亚组Ⅲ的一支,PAN/AtTGA8是花器官发育的调节因子[23-24],也是茎尖分生组织的控制中心,调控茎尖分生组织的大小以及花原基的数目等[25],DE LUIS BALAGUER等[26]和HU等[11]研究还发现,PAN/AtTGA8在拟南芥根尖静止中心细胞中表达,与AtTGA2、AtTGA5和AtTGA6共同调控根的生长发育,在干细胞群动态平衡、细胞氧化还原动态平衡、植株抵抗病菌以及光合作用中共同发挥功能,OsbZIP47在水稻籽粒的粒质量、粒宽的决定中发挥作用[22]。组织表达结果也显示,SiTGA7在枝梗分化期的花序中表达量较高,因此推测,SiTGA7在谷子花器官发育,茎尖、芽尖分生组织调控以及根尖干细胞群维持中起着重要作用,是花器官发育过程中的调节因子,同时还在决定谷子籽粒的粒质量、粒宽的过程中发挥作用。
SiTGA4、SiTGA5、SiTGA9、SiTGA10、SiTGA16与拟南芥TGA家族成员AtTGA2、AtTGA5、AtTGA6、AtTGA9和水稻TGA家族成员OsbZIP03、OsbZIP08、OsbZIP28、OsbZIP63聚在Ⅳ亚组,其中SiTGA5和SiTGA9不含有非编码区。SiTGA5比同一亚组中其他的SiTGAs缺少Motif 10,与同一亚组其他成员主要表达在灌浆期不同,SiTGA5主要表达于未成熟的小穗和籽粒。SiTGA9在灌浆期和未成熟的小穗和籽粒中均有较高表达,而在成熟后籽粒中表达极低。SiTGA10有bZIP_HBP1b-like保守结构域而不含有bZIP superfamily结构域,除在灌浆期根的表达量较高外在成熟后的籽粒中也有较高的表达。拟南芥AtTGA2、AtTGA5、AtTGA6不仅是水杨酸和茉莉酸/乙烯诱导的抗病反应所必需的,而且参与了UV-B胁迫的耐受反应,AtTGA9参与调控拟南芥抗病反应和花器官的发育过程[27-29],水稻中OsbZIP08参与调控水稻的低温抗逆[30],OsbZIP03也在水稻耐盐、耐旱、耐热中发挥作用[22],OsbZIP28在稻瘟病菌侵染应答中发挥重要作用[22],因此推测亚组Ⅳ的谷子TGA家族成员主要参与调控谷子的抗逆反应。
SiTGA14和拟南芥TGA家族的AtTGA1、AtTGA3、AtTGA4、AtTGA7以及水稻的TGA家族基因成员OsbZIP37聚在第Ⅴ亚组,这4个拟南芥成员参与水杨酸调控的植株的抗病反应[15-18],AtTGA1、AtTGA4和AtTGA7还参与了拟南芥花器官的发育过程[31-33],OsbZIP37参与萜类植保素积累来抵御胁迫[22],推测SiTGA14参与谷子中水杨酸调控的抗病反应和花器官的发育。第Ⅵ亚组的SiTGA12主要在灌浆期根中表达,推测其在谷子灌浆期根的生长发育过程中发挥功能。
通过对谷子TGA基因家族的表达模式进行分析,发现SiTGAs在种子萌发期、灌浆期、枝梗分化期以及小穗和籽粒发育时期都有表达,说明谷子TGA家族基因在谷子生长发育中发挥着重要的功能。除了SiTGA3、SiTGA5、SiTGA7以外其他13个基因都在灌浆期根部有很高的表达,说明SiTGAs在谷子根的生长发育过程中发挥重要作用。
本研究通过生物信息学方法对谷子TGA家族基因进行了全基因组鉴定,共鉴定出16个SiTGAs,对这些基因进行了生物信息学分析,结果显示谷子TGA家族基因与高粱TGA家族基因的亲缘关系最近,且亲缘关系相近的SiTGAs其蛋白质理化性质、基因结构和结构域具有保守性。结合TGA家族基因成员的进化关系及SiTGAs在谷子中的表达模式结果可知,谷子TGA家族基因在调控谷子的抗逆反应,花器官发育,茎尖、芽尖分生组织调控以及根尖干细胞维持中起着重要作用。本研究结果为深入研究TGA转录因子在谷子生长发育和胁迫响应过程中的生物学功能提供了参考。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
园林科技(2021年3期)2022-01-19
当代陕西(2021年21期)2022-01-19
中国中医基础医学杂志(2020年1期)2020-03-03
心电与循环(2020年1期)2020-02-27
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
现代农业(2015年5期)2015-02-28
中国全科医学(2015年26期)2015-02-23
