
金桂种子休眠类型鉴定及休眠打破方法探讨
2023-05-15 01:28焦雪辉史喜兵岳长平申潇潇周小娟孙毅宁
山西农业科学 2023年5期
焦雪辉 ,史喜兵 ,岳长平 ,申潇潇 ,周小娟 ,孙毅宁
(1.郑州市农林科学研究所,河南 郑州 450005;2.鄢陵县东华种植农民专业合作社,河南 许昌 461200)
种子休眠的现象普遍存在于高等植物中,林木、果树、花卉、农作物等均存在种子休眠现象。引起休眠的原因包括胚外周组织的机械、物理或化学性抑制等外在原因和胚未分化或发育未成熟、代谢抑制物等内在原因[1]。BASKIN等[2]提出了种子休眠分类系统,该系统主要包括物理休眠、形态休眠、生理休眠、复合休眠和形态生理休眠5个种类。
桂花种子存在休眠现象,一般在4月下旬至5月上旬成熟,通常需要经过6个月以上的常温沙藏才会萌芽。前人对桂花种子休眠进行过研究。张义等[3]研究发现,低温层积能够促进桂花种子发芽。赤霉素浸泡与低温层积相结合效果更好,以3000 mg/L赤霉素浸泡2 d和2000 mg/L赤霉素浸泡2 d或4 d,层积60 d效果最好,发芽率可达62.7%。许岳香[4]研究发现,胚乳的机械作用是造成美国桂花(Osmanthus americanus)种子休眠的主要原因。去种壳+赤霉素处理+赤霉素拌沙低温层积对解除美国桂花种子的休眠最为有效,在处理后45 d能使种子的发芽率达到74%。籽银桂的种皮和胚乳中含有抑制发芽的物质,赤霉素与低温层积结合可以解除种子休眠,其中,以1000 mg/L赤霉素浸种,低温层积105 d的发芽率最高,达72.37%[5]。
目前,对桂花种子休眠的研究多集中在休眠打破方法上,其中,休眠打破多以低温层积和赤霉素浸泡为主,而从形态结构及内源物质等方面对休眠特性进行鉴定,根据休眠特性和原因探索打破休眠方法的研究不多。本试验对金桂(Osmanthus fragransvar.thunbergii)种子种壳吸水性及内源抑制物进行了研究,并通过低温沙藏、种壳机械处理、GA3及硝酸镧浸泡等方法对种子进行了休眠打破,旨在揭示金桂种子的休眠类型,总结休眠打破方法,缩短播种育苗周期,为降低繁育成本提供理论支持。
1 材料和方法
1.1 试验材料
2020年5月初,于郑州市农林科学研究所须水试验场采集成熟的金桂果实,去除果肉,洗净晾干,从中挑选大小均匀、饱满、无病虫害的种子,用3%高锰酸钾溶液浸泡20 min,蒸馏水冲洗3次,放置于实验室备用。
试验中GA3购自北京中科起源科技有限公司,硝酸镧购自阿拉丁生化科技股份有限公司。
1.2 试验方法
1.2.1 休眠类型鉴定试验
1.2.1.1 种壳吸水性试验 分别取去壳种子及划口种子(从胚一端开始,用解剖刀向下对称划开种壳,至种子1/2处,露出胚但不损伤胚)各20粒,以完整种子作为对照。种子称量初始质量(G0),放入锥形瓶中,加入30 mL蒸馏水,置于25 ℃恒温环境中,每隔一定时间取出种子用吸水纸吸干表面水分后称质量(Gi),重复3次,直至种子质量不再增加为止。前8 h,每浸泡2 h取一次称质量,之后每隔12~24 h取一次。以净吸水量随时间的变化来表示种子的吸水率(Vi),并绘制吸水曲线。
1.2.1.2 金桂种子内源抑制物对白菜籽发芽的影响 取新鲜的种子,剥开种子并分离种壳和胚乳,分别称取1 g,放入预冷的研钵内研碎,加入80%甲醇溶液20 mL,摇晃均匀,放入5 ℃冰箱恒温浸提24 h,每4 h晃动一次,24 h后过滤,连续浸提3次,将3次滤液合并后倒入旋转蒸发仪,减压蒸干后用蒸馏水冲洗,最后定容到100 mL作为种壳和胚乳浸提液。重复3次。
挑选饱满的白菜籽,用0.5%高锰酸钾溶液消毒,蒸馏水冲洗净后自然风干备用。将培养皿中放入2层滤纸,加入浸提液20 mL,放入白菜籽30粒,培养皿上覆一层保鲜膜,均匀扎孔保证透气,放入25 ℃恒温光照培养箱培养,期间保持滤纸湿润。以蒸馏水培养作为对照。每个处理重复3次。48 h后统计每个培养皿中发芽的白菜籽数量,以发芽长度超过种子直径为标准,计算平均发芽率;72 h后测量根长,记录白菜籽生长情况。
1.2.2 休眠打破试验
1.2.2.1 GA3质量浓度对金桂种子发芽的影响 取划口种子30粒,分别放入质量浓度为1000、2000、3000 mg/L的GA3溶液中浸泡48 h,以蒸馏水浸泡作对照。浸泡后用蒸馏水洗净,放入陶盆中5 ℃低温沙藏处理60 d后取出种子,用3%高锰酸钾溶液浸泡20 min,蒸馏水冲洗3次,放入25 ℃光照培养箱培养,光照16 h、黑暗8 h。每组重复3次,连续5 d不再发芽时试验结束,以发芽长度超过种子直径为标准统计发芽率。
1.2.2.2 硝酸镧质量浓度对金桂种子发芽的影响取划口种子30粒,分别放入质量浓度为10、20、30 mg/L的硝酸镧溶液中浸泡48 h,以蒸馏水浸泡作对照。浸泡后用蒸馏水洗净,放入陶盆中5 ℃低温沙藏处理60 d后取出种子,用3%高锰酸钾溶液浸泡20 min,蒸馏水冲洗3次,放入25 ℃光照培养箱培养,光照16 h、黑暗8 h。每组重复3次,连续5 d不再发芽时试验结束,统计发芽率。
1.2.2.3 种壳处理方式对金桂种子发芽的影响 分别取完整种子、划口种子、去壳种子各30粒,用1000 mg/L GA3溶液浸泡48 h,蒸馏水冲洗干净后,放入陶盆中5 ℃沙藏60 d,取出种子,用3%高锰酸钾溶液浸泡20 min,蒸馏水冲洗3次,放入25 ℃光照培养箱培养,光照16 h、黑暗8 h。每组重复3次,连续5 d不再发芽时试验结束,统计发芽率。
1.2.2.4 低温沙藏时间对金桂种子发芽的影响取划口种子30粒,用1000 mg/L GA3溶液浸泡48 h,蒸馏水冲洗干净后,分别放入陶盆中5 ℃沙藏30、60、90 d,以沙藏0 d作对照。种子沙藏后用3%高锰酸钾溶液浸泡20 min,蒸馏水冲洗3次,放入25 ℃光照培养箱培养,光照16 h、黑暗8 h。每组重复3次,连续5 d不再发芽时试验结束,统计发芽率。
1.2.3 综合因素对金桂种子发芽的影响 在单因素试验基础上,选出单因素中对休眠打破效果较好的处理进行优化,综合几个因素进行种子休眠打破试验(表1)。种子在不同溶液中的浸泡时间均为48 h,消毒、培养、发芽率统计等处理同单因素试验,每处理取30粒种子,重复3次。

表1 多因素处理方法Tab.1 Multifactor treatment method
1.3 数据分析
试验采用Excel进行数据统计,SPSS 23.0进行数据分析。
2 结果与分析
2.1 金桂种子休眠类型鉴定
金桂种子吸水曲线如图1所示。

图1 金桂种子吸水曲线Fig.1 Water absorption curve of Osmanthus fragrans var. thunbergii seeds
从图1可以看出,去壳种子和划口种子的最终吸水率均显著高于完整种子。完整种子吸水率最终为26.73%,去壳种子的吸水率为37.46%,划口种子的吸水率为35.33%。划口种子和去壳种子前6 h为快速吸水期,吸水率迅速上升,8 h后吸水率减缓,3 d后完整种子吸水达到饱和,6 d后划口种子吸水达到饱和。去壳种子在0~12 h,种子迅速吸水,12~48 h种子吸水率上升速度最快,6 d后吸水达到饱和。由此可以看出,金桂的种壳对种子吸水存在一定的阻碍作用。
由表2可知,培养24 h后,3种培养条件下白菜籽发芽率差异显著,蒸馏水培养24 h后白菜籽发芽率可达到100%,种壳和胚乳浸提液培养的白菜籽发芽率均显著低于对照(P<0.05),且胚乳浸提液培养的白菜籽发芽率显著低于种壳浸提液(P<0.05)。48 h后白菜籽发芽率差异仍然显著,规律与24 h时相同,种壳和胚乳浸提液培养条件下白菜籽发芽率有所升高,胚乳仍然最低,仅为57.78%。48 h后3种培养条件下根长差异显著,蒸馏水培养条件下根长最长,达到23.50 mm,胚乳浸提液培养条件下根长最短,仅为3.47 mm。72 h后 3种培养液对白菜籽根长的影响依然差异显著。蒸馏水培养的白菜籽根长达到47.43 mm,比48 h时增加了23.93 mm;种壳浸提液培养的白菜籽根长也有所增加,胚乳浸提液依然最短,且仅比48 h时增加了0.06 mm。由此可见,种壳和胚乳浸提液对白菜籽的萌发及根的生长有明显的抑制作用,其中,胚乳的抑制作用更明显,说明种 壳及胚乳中可能含有抑制种子发芽的物质。

表2 白菜籽发芽情况Tab.2 Germination of Chinese cabbage seeds
2.2 单因素处理对金桂种子休眠打破的影响
在所有处理中,金桂种子在放入培养箱第2天即开始发芽,培养15 d左右不再有新种子发芽。所有处理对种子开始发芽时间的影响差异不大。
由图2可知,GA3质量浓度为1000、2000 mg/L时,种子的发芽率显著高于GA33000 mg/L,也显著高于对照(P<0.05),1000、2000 mg/L GA3间发芽率差异不显著。其中,GA3质量浓度为1000 mg/L时种子发芽率最高,可达51.11%,而GA3质量浓度为3000 mg/L时种子发芽率仅为15.56%,与对照间差异不显著。可见,高质量浓度的GA3可能对种子发芽有抑制作用。

图2 GA3质量浓度对休眠打破的影响Fig.2 Effect of GA3 mass concentration on dormancy breaking
硝酸镧质量浓度对金桂种子发芽率影响差异显著(P<0.05),3个质量浓度条件下种子发芽率均显著高于对照。其中,硝酸镧30 mg/L时发芽率最高,且显著高于其他2个质量浓度,种子发芽率可达44.44%。硝酸镧质量浓度为20 mg/L时,种子发芽率为37.78%,显著高于质量浓度为10 mg/L时的27.78%(图3)。可见,硝酸镧对种子发芽有促进作用,高质量浓度的硝酸镧对种子发芽促进作用更明显。

图3 硝酸镧质量浓度对休眠打破的影响Fig.3 Effect of lanthanum nitrate mass concentration on dormancy breaking
将金桂种子进行去壳和划口处理后,种子发芽率显著提高,其中,划口种子发芽率最高,达到41.11%,与去壳种子发芽率间差异不显著。完整种子发芽率仅为11.11%,显著低于其他2种处理(图4)。可见,种壳对种子发芽存在一定的机械阻碍。在试验中观察到,去壳种子尽管完全摆脱了种壳的机械阻碍,但其发芽率却并非是最高的,可能是因为种子完全去壳后,胚乳直接暴露在空气中,更容易被细菌污染,从而导致发芽率降低。

图4 种子处理方式对休眠打破的影响Fig.4 Effect of seed treatment on dormancy breaking
在不同贮藏时间对金桂种子发芽影响试验中,贮藏90 d后种子的发芽率最高,达到52.22%,与贮藏60 d间差异不显著,但二者均显著高于贮藏30 d和对照的发芽率(P<0.05)。贮藏30 d后种子发芽率为16.67%,而对照的种子发芽率仅为5.56%(图5)。可见,低温贮藏时间对种子发芽影响显著,贮藏时间越长,种子发芽率越高。
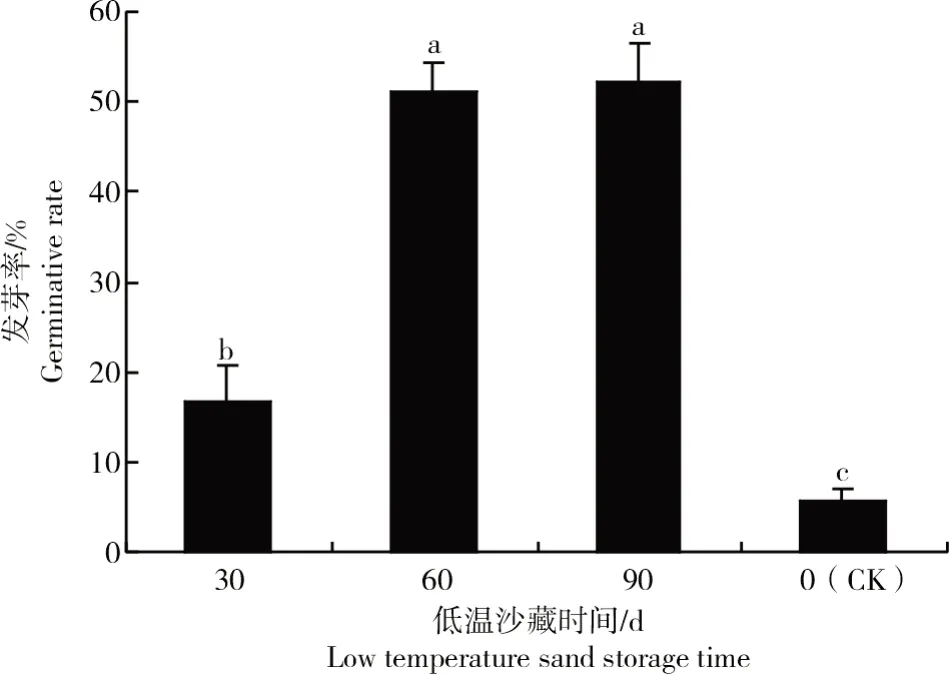
图5 低温沙藏时间对休眠打破的影响Fig.5 Effect of low temperature sand storage time on dormancy breaking
从图2~5还可以发现,低温沙藏时间对种子休眠打破的影响最显著,低温沙藏90 d与未经过低温沙藏的种子萌发率相差46.66百分点,而种壳处理方式中划口种子与完整种子萌发率相差30百分点,可见,生理后熟可能是造成金桂种子休眠的主要原因,而种壳的机械阻碍可能是造成种子休眠的次要原因。
2.3 多因素处理对金桂种子休眠打破的影响
因单因素处理中金桂种子发芽率最高也仅能达到52.22%(图5),为进一步提高种子发芽率,对单因素试验进行优化,综合几个因素对金桂种子休眠打破进行试验。从图6可以看出,所有处理条件下种子发芽率均显著高于对照,其中,处理1、4、5、7条件下种子发芽率均在80%以上。处理5条件下种子萌发率最高,可达87.78%,与处理1、4间差异不显著,说明在其他因素一致的情况下,硝酸镧质量浓度分别为40、50 mg/L时,与30 mg/L时对发芽率影响无显著差异。在同样浓度下,同时使用GA3和硝酸镧处理的发芽率要远远高于单独使用其中一种时的发芽率。处理1条件下种子发芽率显著高于处理2、3,而处理2、3之间差异不显著,说明GA3质量浓度为1000 mg/L时休眠打破效果要远远好于低质量浓度。处理6条件下发芽率要显著低于处理1,说明相同条件下划口种子休眠打破效果要显著好于去壳种子。处理1条件下种子发芽率与处理7间差异不显著,但二者均显著高于处理8,说明5 ℃沙藏60、50 d对种子发芽率影响差异不大,但显著高于40 d时发芽率,由此可见,金桂种子在低温沙藏50 d后可能已经解除休眠了。

图6 多因素处理对种子休眠打破的影响Fig.6 Effect of multifactor treatment on seed dormancy breaking
3 结论与讨论
造成种子休眠的因素很多,主要包括环境因素,如温度[6-7]、光照[8]等,以及种子内源因素,如种皮结构[9]、内源抑制物质[10]、种胚发育不成熟[11]、胚乳的机械阻隔[12-13]等。打破种子休眠常用的方法有种皮去除、层积处理、外源激素及化学试剂处理等,其中,外源激素打破种子休眠具有高效、便捷等优点[14]。杨秀莲等[15]对紫梗籽银桂种子休眠原因进行了初步探讨,认为种皮和种仁中的抑制物质可能是引起种子休眠的原因,而坚硬的外种皮并不是引起休眠的原因。本研究中对金桂种壳的吸水性和内源抑制物进行了初步研究,结果发现,去掉坚硬的种壳或者在种壳上划口处理,最终的吸水率要高于完整种子,说明金桂种子的种壳对吸水性存在一定的阻碍作用。但金桂种子的种壳比较薄,即使完整种子吸水速率也并非特别低,可以保证种子萌发对水分的需求,因此,种壳的机械阻碍并不是造成金桂种子休眠的主要原因。
种子的内源抑制物质种类具有多样性,主要包括辛酸、棕榈酸[16-17]、酚类、脂类、酮类[18]等,这些物质对种子萌发都有抑制作用,但作用方式和机制各不相同。施婷婷等[19]对紫梗籽银桂果皮、种实内源抑制物进行了提取和鉴定,发现存在多种抑制物质,主要为脂肪酸类物质。本研究中利用白菜籽萌发试验,间接证明了金桂种子中也存在抑制物质,主要抑制白菜籽根的发芽和伸长,且胚乳的抑制作用要比种壳明显。不过,本研究所得出的抑制作用是针对白菜籽的,且浸提液未进行稀释,浓度较高,金桂种子的内源抑制物质对其自身种子萌发是否存在抑制作用,以及抑制物浓度达到多少才会对其种子萌发产生抑制作用,还需要进一步论证。
种子由抑制物质起主要作用转向促进物质占主导地位,是解除休眠的决定性因素[20]。休眠解除过程受多种激素控制。ABA含量与GA3/ABA值可能是解除休眠的关键因素[21]。种子经过层积处理后,可以减少萌发抑制物,调控种子内部激素水平向利于休眠解除的方向发展,同时能够增加种皮的透性,促进种子生理后熟。杨秀莲等[5]研究认为,赤霉素结合低温层积可以有效解除桂花种子的休眠。袁王俊等[22]则研究认为,使用赤霉素和3个月层积处理不能促进桂花种子萌发,但层积3个月后进行离体培养,再在培养基中加入NAA和BA可以打破休眠,促进桂花种子萌发。本研究分别对赤霉素和低温层积时间进行了梯度试验,结果表明,GA3质量浓度为1000、2000 mg/L均可以提高金桂种子发芽率,且显著高于对照和GA3质量浓度为3000 mg/L时的种子发芽率,这与杨秀莲等[15]研究结论基本一致。低温层积60、90 d均可以显著提高种子发芽率,且显著高于对照和低温层积30 d时的发芽率,这与杨秀莲等[15]研究结论不一致。一方面可能是试验材料本身的差异,另一方面可能是因为层积温度和层积容器的差异,杨秀莲等[15]试验中所用材料为紫梗籽银桂,在塑料棚中2~4 ℃低温层积,而本研究中用的是金桂,在陶盆中5 ℃低温层积。通过试验还发现,GA3和低温层积处理对种子发芽的促进作用要显著强于种壳机械处理,因此,可以断定金桂种子应该属于复合休眠类型,同时存在生理休眠与物理休眠。生理后熟是导致桂花种子休眠的主要因素,而种壳的机械阻碍只是次要因素。
许多研究和综述[23]已经指出,稀土元素可促进植物生长发育,提高作物产量,提高作物淀粉酶、蛋白酶、脂肪酶、过氧化物酶、硝酸还原酶和酯酶等活性及土壤酶的活性,促进新陈代谢,说明稀土是一种新型的植物生长调节剂。镧是17种稀土元素中最重要和最活泼的元素[24]。本研究利用硝酸镧对金桂种子进行休眠打破试验,发现其对金桂种子休眠的打破有促进作用,可能是因为稀土元素镧对种子萌发的促进作用,镧能够提高种子或种芽中与生长发育相关的酶活性,从而加快大分子物质的代谢,提供种子、种芽萌发所需能量和养分,因此,促进了种子、种芽的萌发。而将硝酸镧与赤霉素混合,同时结合低温层积和种壳机械处理,可以进一步提早解除休眠,提高种子发芽率。GA31000 mg/L+硝酸镧50 mg/L+5 ℃沙藏60 d+种子划口处理组合对金桂种子休眠打破效果最好。
猜你喜欢
防护林科技(2020年11期)2020-12-30
作文小学中年级(2020年9期)2020-09-29
河北林业科技(2020年3期)2020-03-23
种子(2019年7期)2019-08-13
财讯(2019年33期)2019-08-04
中国食品学报(2019年3期)2019-01-13
甘肃农业科技(2018年3期)2018-05-18
作文周刊·高二版(2016年32期)2017-04-13
蔬菜(2016年10期)2016-03-27
武夷学院学报(2015年3期)2015-07-18
