
新疆野生樱桃李PsoRPM2与PsoLFY互作调控植株开花的分子机制
2023-11-09 14:19高朝远关平印蒲文江袁梓珍罗嗣芳胡建芳
中国农业大学学报 2023年11期
高朝远 关平印 蒲文江 袁梓珍 罗嗣芳 胡建芳
(中国农业大学 园艺学院,北京 100193)
高等植物开花是其生命活动中的重要阶段,也是由营养生长向生殖生长转变的重要标志[1]。植物只有在适宜的时期开花才能完成授粉受精形成种子,繁衍后代并确保物种的自然延续[2]。早开花可以在农业生产中促进早坐果、早成熟和早上市从而提高经济效益[3-4]。另外对于多年生实生繁殖的果树而言,由于存在着较长的童期,提早开花可以缩短育种周期,提高育种效率,节约土地,获得早期丰产[5-6]。因此,研究植物的早花对于园艺植物具有重要意义。
植物的开花过程涉及到成花诱导、花发育和花器官形成等阶段,每一个发育阶段都必须在前一个阶段完成的基础上而展开[7]。目前大多数研究都集中在成花诱导方面,因为这一阶段容易受到外界环境和植物内部信号共同影响,是比较敏感的时期,也是调节植物开花时间和数量的关键时期[1]。对拟南芥的研究表明,成花诱导主要涉及6条途径[8],即春化途径、光周期途径、赤霉素途径、自主途径、温敏途径和年龄途径。这些途径彼此独立又相互交联构成复杂而精密的调控网络[9-10]。虽然模式植物的成花途径已经相当清晰,但对于多年生果树而言其成花途径有别于一、二年生植物。因为果树一旦度过童期或营养生长阶段每一年都循环往复地进行花芽分化、休眠、萌芽、开花坐果和果实成熟等,并且果树的花芽分化时间较长,一般需要跨越2个年度,因此果树的成花调控途径可能更为复杂。但目前对多年生果树的成花调控途径还缺乏系统的研究。
成花调控网是由自身遗传因素和环境因素共同决定的复杂过程,其内在原因是通过一系列成花基因在时间和空间上按特定顺序进行特异表达从而精准调控植物的开花[11]。目前对拟南芥6条成花途径中不同成花基因的表达顺序和功能都有较系统和清晰的研究。在这些途径中存在着一些成花信号整合因子,如CONSTANS(CO)、FlOWERINGLOCUST(FT)、SUPPRESSOROFCONSTANS1(SOC1)、LEAFY(LFY)、SQUAMOSAPROMOTERBINDINGPROTEIN-LIKE(SPL)、TERMINALFLOWER1(TFL1)和FLOWERINGLOCUSC(FLC)等[12],它们对植物开花起着关键作用。在这些基因中既有促进成花的基因也有抑制成花的基因。对拟南芥的研究表明,至少有180个以上的基因可能参与了开花调控的过程[8]。但由于果树在花芽分化中经历的内外环境复杂而多变,加之果树遗传转化较为困难,耗时长,不宜重复验证,对于成花基因的研究大多参照模式植物进行,多数成花基因在果树中的具体功能还不清楚。
植物的开花除了受到成花基因的调控,还受到寒冷、干旱、盐分和病原体感染等环境胁迫影响[12-15]。拟南芥中的核因子Y(Nuclear factor Y)包括NF-YB2和NF-YB3,与胁迫应答有关,通常它们需要形成同源或异源二聚体或三聚体才能发挥作用。近期的研究发现,CO可以与NF-Y互作[16-17],CO可以代替NF-YA与NF-YB、NF-YC形成三聚复合物,YB2与YB3通过招募CO结合到FT启动子上,促进植物开花[18-19]。这表明植物的开花调控还存在着一些未知的领域。
新疆野生樱桃李(Prunussogdiana)是原产中国西部天山山脉原始森林中的一种珍贵野生果树[21],具有耐瘠薄土壤、抗盐碱、耐湿、抗寒性强和抗病等特点[21-25]。与杏、梨、桃和樱桃具有嫁接亲和性,可以作为核果类果树的优良砧木[26-27]。前期课题组已从新疆野生樱桃李实生抗病单株中克隆得到psoRPM2抗南方根结线虫基因,并通过遗传转化鉴定了其功能;同时发现转psoRPM2基因烟草大多表现为早花[28]。本研究在此基础上,观察了新疆野生樱桃李实生群体的开花类型和花芽分化特点,分析了新疆野生樱桃李不同开花类型的关键成花基因表达,对转psoRPM2基因烟草的开花特性和与早花相关的成花基因进行了研究,同时探讨了psoRPM2与PsoLFY蛋白之间的关系,阐明了PsoRPM2基因引起的早花现象,以期明确psoRPM2调控新疆野生樱桃李开花时期的作用机制。
1 材料与方法
1.1 材料
利用定植于中国农业大学上庄实验站的新疆野生樱桃李早花与晚花植株,于2021年6月15号、7月15号、8月15号、9月15号和10月15号分别收集花芽与叶芽。一部分用FAA固定,用于石蜡切片的制备;另一部分置于-80 ℃保存用于RNA提取。转psoRPM2基因烟草利用实验室前期的组培苗。
1.2 试验方法
1.2.1开花生物学性状及解剖与组织结构观察
开花生物学性状是利用本实验室常年记载的开花物候期数据,通过统计分析,确定不同单株的开花特性。同时利用解剖刀对不同发育时期的花芽进行徒手切片,置于体试镜下观察。将固定液(FAA)固定的样品经过不同浓度的乙醇脱水、二甲苯透明处理、沉浸石蜡和包埋等常规石蜡切片制作过程,获得含有不同试验材料的蜡块;在切片机上切成8~12 μm厚的切片,用番红-固绿染色后封片,在OlympusBX-51光学显微镜观察和拍照。每个发育时期的花芽各统计20个样品。
1.2.2RNA提取、半定量PCR及实时荧光定量PCR
采用CTAB法提取新疆野生樱桃李花芽和叶芽的总RNA,通过反转录试剂盒(Promega)反转成cDNA,用于半定量RT-PCR和实时荧光定量表达(RT-qPCR)分析。根据同源克隆得到的PsoLFY、PsoFT、PsoTFL、PsoFLC、psoRPM2基因序列和在NCBI上查询到的烟草NtLFY(XM_016593842.1)、NtFT(KC485967.1)、NtTFL(XM_016590731.1)、NtFLC(XM_016599652.1)、NtSOC(NM_001326029.1)、NtSPL(NM_001326256.1)、NtFD(XM_016613899.1)基因序列设计RT-PCR和RT-qPCR引物(表1),RT-PCR反应产物用1.5%的琼脂糖凝胶电泳检测。RT-qPCR反应体系使用Applied Biosystems 7500(Thermo Fisher Scientific,40 ℃)进行40个循环,95 ℃预变性10 s和60 ℃变性30 s。使用2-ΔΔCt方法(Livak和Schmittgen,2001)检测mRNA的表达水平,使用RPⅡ作为参照基因的标准。用Excel整理数据,用GraphPad Prism 5软件进行作图。每个发育时期的花芽和叶芽器官至少3个生物学重复。

表1 本研究所用引物序列及退火温度
1.2.3基因克隆与酵母双杂交
利用上述得到的樱桃李cDNA模板,分别克隆PsoLFY和PsoRPM2基因,将PsoLFY和PsoRPM2分别构建至酵母双杂pGBKT7和pGAKT7质粒上。将BD-PsoLFY与AD-PsoRPM2、阳性对照、BD-empty与AD-PsoRPM2、BD-PsoLFY 与AD-empty、BD-empty与AD-empty转入酵母感受态细胞,涂布于SD/-Trp/-Leu二缺培养基平板上,30 ℃培养箱中倒置培养4~5 d;挑选6个单菌落于SD/-Trp-Leu-His三缺培养基上,倒置培养3~5 d;将阳性克隆再点于SD/-Trp-Leu-His-Ade四缺培养基;对生长出来的菌落进行X-α-Gal染色(4 mg/mL,每个菌落5 μL),通过显色分析α-半乳糖苷酶活性,观察并拍照。
1.3 数据处理
采用Excel对新疆野生樱桃李花芽切片原始数据进行统计分析,使用Graphpad prism 6作图。
2 结果与分析
2.1 新疆野生樱桃李开花生物学性状
本研究对定植于上庄实验站的112株实生野生樱桃李开花生物学性状进行观察,开花类型分为极早花、早花、中花、晚花和极晚花5种类型。2021-03-25拍照发现极早花类型中已有30%以上开始露白,早花类型中有10%~30%露白,中花类型中露白比率不足5%,而晚花类型中花芽刚开始膨大,极晚花类型中大多数花芽还未有膨大(图1(a))。对112株实生野生樱桃李不同开花类型统计表明,极早花、早花、中花、晚花和极晚花类型的比率分别为11%、24%、44%、18%和3%(图1(b))。极早花与极晚花类型的开花时期平均相差10~14 d。

(a)同一时期极早花、早花、中花、晚花、极晚花的发育状态;(b)实生群体中不同开花类型所占的比例;(c)同一发育时期早花和晚花不同发育状态的统计分析。条形图上的小花分别为不同发育时期的代表图。
本研究从早花和晚花类型中分别选取2株树体在开花前的未露白期(平均花蕾直径(4±1) mm)、露白期(平均花蕾直径(5±1) mm)、小蕾期(平均花蕾直径(6±1) mm)和大蕾期(平均花蕾直径8±1 mm)进行统计分析。结果显示早花类型的未露白期、露白期、小蕾期和大蕾期比率分别为2.20%、7.50%、5.20%和85.10%,而晚花类型的比例则为19.60%、9.50%、8.30%和62.60%(图1(c))。
2.2 不同开花类型早花和晚花植株的花芽分化过程
由于新疆野生樱桃李实生群体存在早开花和晚花类型,本研究对早花和晚花类型花芽分化过程的外观形态和组织解剖学形态进行了观察。利用徒手切片在6月15号(大约是上一年分化的花芽开花后65 d)分别观察早花和晚花的花芽形态。此时早花和晚花都已分化出来5~7层鳞片,早花植株的生长点已经隆起呈半圆状,晚花植株的生长点还没有明显的隆起(图2(a))。7月15号早花植株的花芽生长点已经呈现柱状,并已完成萼片的分化;此时晚花植株的花芽生长点才明显隆起(图2(b))。到8月15号早花植株已经进入到花瓣原基分化期,而此时晚花植株完成了萼片的分化,还未进行花瓣原基的分化(图2(c))。9月15号早花植株已经进入到雄蕊原基分化期,此时的晚花植株刚进入花瓣原基分化期(图2(d))。到入冬前的10月15号早花植株已经进入雌蕊分化期,晚花植株完成了雄蕊分化期(图2(e))。早花和晚花植株的开花状态存在明显不同(图2(f))。

图2 新疆野生樱桃李早花与晚花植株花芽外部及解剖结构
进一步利用石蜡切片观察花芽分化过程,发现早花类型和晚花类型的花芽分化过程并没有明显差异,都经历了未分化时期、原基膨大期、萼片原基、花瓣原基、雄蕊原基和雌蕊原基分化期。在未分化期茎端生长点较小,呈圆锥形,细胞排列整齐,鳞片紧紧包围在生长点外部(图3(a));其后分生细胞膨大,生长点开始突起,生长点变大,此时外部的芽鳞片向外扩展变得疏松(图3(b));随着生长点的继续变宽变大,突起明显呈现半圆形(图3(c));之后生长点逐渐变长、变宽、顶端扁而平,在生长点外围开始凸起即为萼片原基(图3(d));随着萼片原基的不断向内弯曲分化,在伸长的萼片原基内侧产生新一轮凸起即为花瓣原基(图3(e));在花瓣原基内侧继续分化形成新的凸起分化出雄蕊原基(图3(f));其后雄蕊原基继续分化(图3(g));随后分化雌蕊原基(图3(h));分化完全的花芽包括鳞片、花萼、花瓣、雄蕊和雌蕊原基(图3(i)和(j));随后在雌蕊原基中央开始分化心皮边缘组织(图3(k));进一步心皮边缘组织融合形成单心皮子房(图3(l))。
根据对早花和晚花类型进行统计分析发现,6月15日早花和晚花类型花芽大多数处于未分化阶段,其比率分别为89%和95%。7月15日早花类型有70%处于分化初期,而晚花类型的70%处于分化初期。到8月15日早花类型有60%处于花瓣原基分化期,并且有30%的花芽已经进入雄蕊原基分化期;此时晚花类型的60%花芽处于萼片原基分化期,有10%花芽处于花瓣原基分化期,没有花芽进入雄蕊原基分化期。9月15日早花类型的70%处于雄蕊原基分化期,有15%的花芽进入雌蕊原基分化期;晚花类型的70%花芽处于花瓣原基分化期,23%的花芽进入雄蕊原基分化期,没有雌蕊原基分化期的花芽。到10月15日早花类型已经有90%的花芽完成了雌蕊原基的分化,而晚花类型只有60%的花芽完成雌蕊原基的分化(表2)。
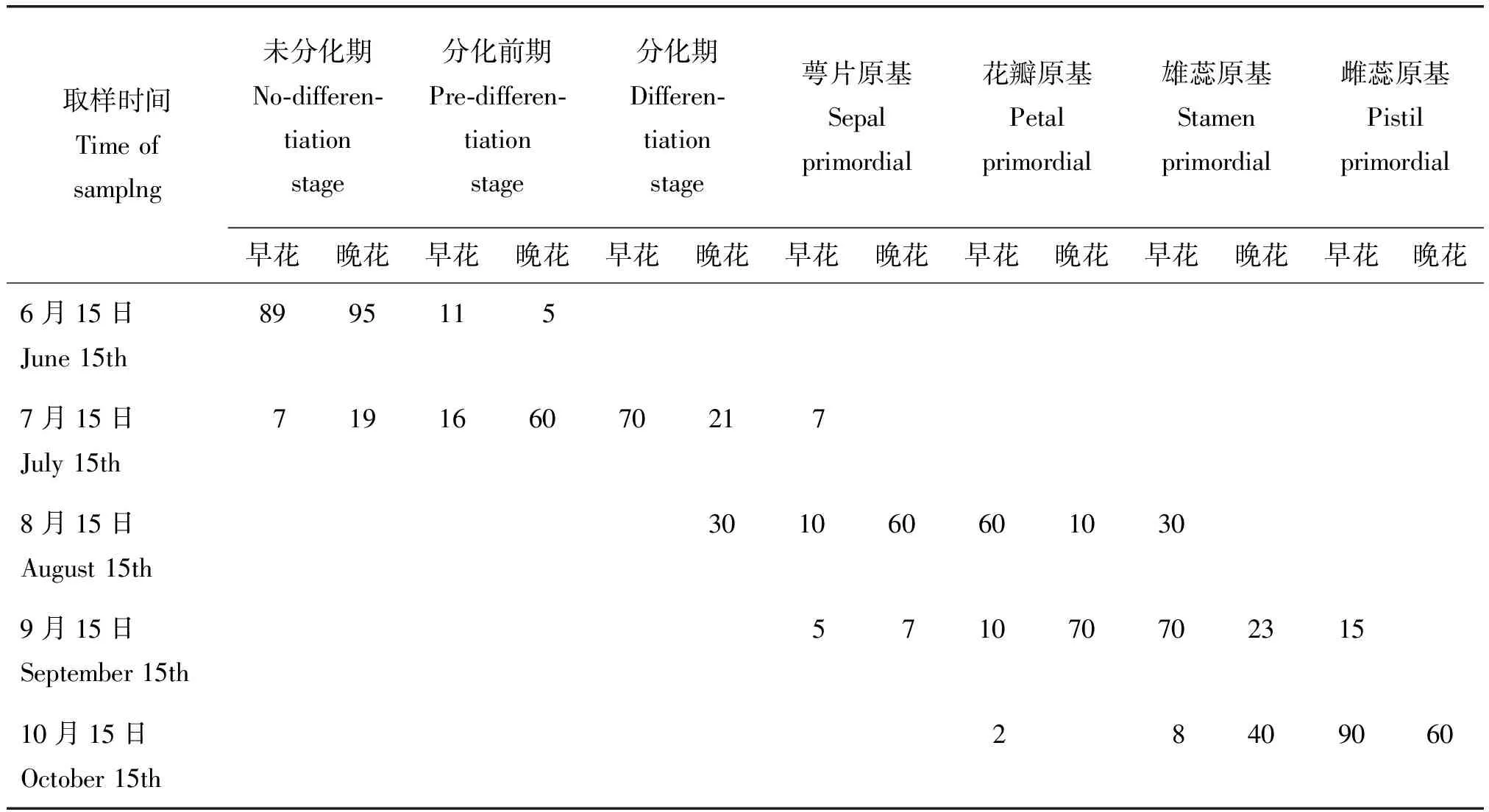
表2 不同发育时期早花和晚花类型花芽分化所占比例
2.3 相关成花基因在野生樱桃李早花和晚花花芽中的表达
对早花和晚花花芽分化过程中相关成花基因的表达进行分析,结果显示,PsoLFY基因在7月15日和8月15日的早花花芽中其表达高于晚花,但在9月15日和10月15日的早花花芽中其表达量却低于晚花类型(图4(a))。PsoFT基因表达与PsoLFY基因相似,但在7月15日和8月15日的早花花芽中的PsoFT与晚花相比差距更大(图4(b))。PsoFLC基因在7月15日的早花花芽中表达量高于晚花类型,其余的8月15日、9月15日和10月15日PsoFLC基因的表达都是晚花高于早花类型(图4(c))。PsoTFL基因表达在早花与晚花花芽中差别不大(图4(d))。
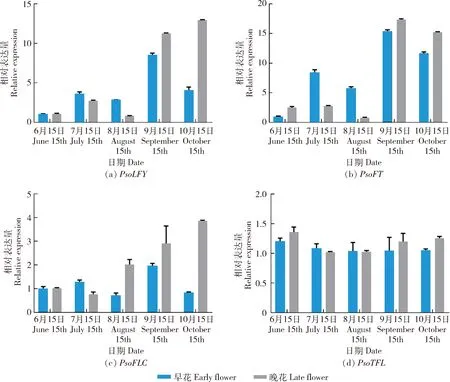
误差线表示标准误(SE)。数值为平均值±标准误。
2.4 转PsoRPM2基因烟草可以提早开花
本研究将PsoRPM2基因转入烟草导致出现早花现象,共得到5个转基因植系。对转PsoRPM2基因烟草的早花性状进行观察发现,其中有1、2和5株系表现为早花,从愈伤再生到植株开花需要的天数分别为64、59和57 d,平均开花天数为60 d。而普通烟草在相同环境条件(4 000 LX光照强度、16 h光照时间、25 ℃)下生长1年左右时间才能开花(图5(a))。转基因株系3和4未表现为早花。另外3个转PsoRPM2基因烟草在株高、叶片数和节间数方面都比同一时期的对照要多,尤其是花序数明显高于对照(图5(c))。
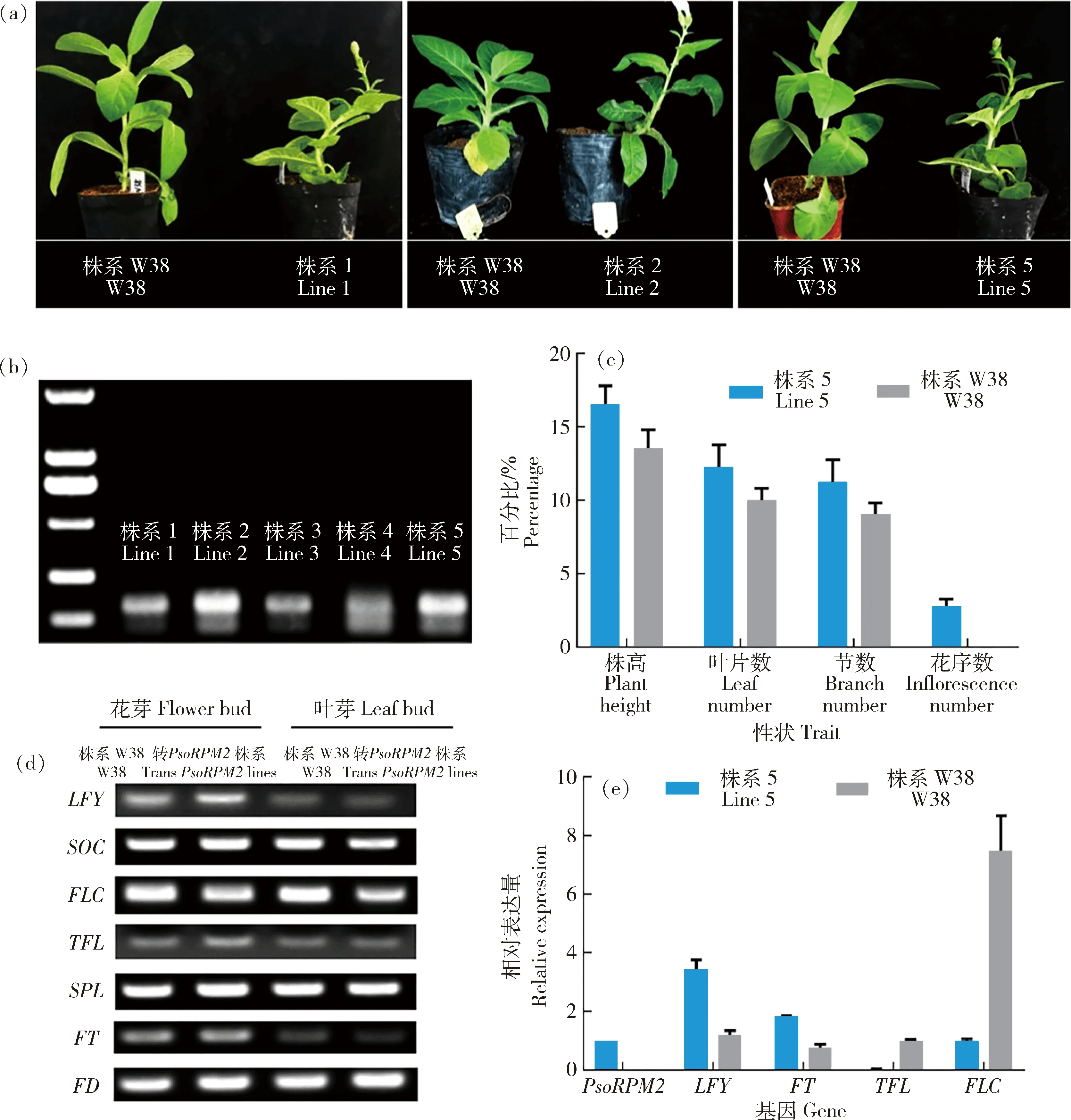
(a)野生型和转PsoRPM2基因烟草表型图;(b)PsoRPM2基因在不同转基因烟草株系中的表达情况;(c)野生型和转PsoRPM2基因烟草株系5生理性状统计图;(d)RT-PCR分析开花相关基因在野生型和转PsoRPM2基因烟草株系5花芽和叶芽中的表达情况;(e)RT-qPCR分析开花相关基因及PsoRPM2基因在野生型和转PsoRPM2基因烟草株系5花芽中的表达情况。株系W38为野生型烟草,株系1、2和5分别为转基因烟草。
对转PsoRPM2基因烟草花芽和叶芽中的相关成花基因进行RT-PCR分析。结果显示,在叶芽中所有成花基因在转PsoRPM2基因和对照之间没有差异;但在花芽中,PsoLFY和PsoTFL基因在转PsoRPM2基因和对照之间存在差异,而PsoSOC、PsoFLC、PsoSPL、PsoFT和PsoFD基因在两者之间都没有明显差异(图5(d))。进一步通过RT-qPCR分析转基因株系3花芽中的成花基因表达,发现PsoLFY和PsoFT基因在转PsoRPM2基因烟草花芽中的表达明显高于对照,而PsoFLC和PsoTFL基因表达则明显低于对照(图5(e))。
2.5 野生樱桃李早花和晚花类型芽中的PsoRPM2基因表达
上述研究表明转PsoRPM2基因烟草可以促进植物提前开花,本研究进一步分析野生樱桃李早花和晚花花芽与叶芽中的PsoRPM2基因表达。结果显示,在6月15日PsoRPM2基因的表达量最低,同时在早花和晚花类型的花芽中其表达量几乎没有差异。到7月15日早花类型花芽中的PsoRPM2基因表达明显高于晚花类型,两者之间的差距最大。
在8月15日早花类型略高于晚花类型,但到9月15日和10月15日时早花和晚花类型花芽中的PsoRPM2基因表达几乎没有差异(图6(a))。在早花和晚花类型的叶芽中PsoRPM2基因表达没有明显差异(图6(b))。
2.6 野生樱桃李PsoLFY与PsoRPM2的酵母双杂分析
由于转PsoRPM2基因株系花芽中的LFY基因表达明显高于对照,同时在新疆野生樱桃李7月15日成花诱导的花芽中PsoRPM2基因表达明显高于晚花,本研究认为转PsoRPM2基因植株之所以早花可能与LFY基因表达有关。因此进行了酵母双杂试验,结果显示PsoRPM2与PsoLFY可以进行互作(图7)。
3 讨论与结论
野生资源是获得果树重要抗逆性状的主要来源,是人们选育新品种开展生物技术研究的主要基因来源[29]。新疆樱桃李原产于天山野果林,在中国仅分布在新疆伊犁地区霍城县境内婆罗科努山的大西沟和小西沟,这一地区属于温带荒漠区,野生果树资源十分丰富,是栽培苹果、杏、核桃、欧洲李和樱桃李的起源中心,对研究果树的起源、进化和演变具有重要意义[30-31]。在自然界,野生樱桃李主要通过自然实生繁衍后代,形成了植株性状、果实形状、颜色、枝条和花等特征多种多样的群体[32-33]。本研究通过对新疆野生樱桃李物候期的观察,发现其开花时间存在着较大差别,早开花和晚开花的类型开花时间大约相差14 d,表明新疆野生樱桃李开花生物学具有多样性。同时在植株高矮、枝条生长与节间长短、花形态、果实颜色和大小等方面也存在着较大差异[34]。
植物开花时间与花芽分化密切相关,花芽分化的进程是决定开花时间的主要因素[35]。从新疆野生樱桃李花芽分化的阶段来看,早开花与晚开花类型之间没有明显差异,两者都经历了未分化期、分化前期、分化初期、萼片原基分化期、花瓣原基分化期、雄蕊原基分化期和雌蕊原基分化期,这一结果与李会芳等[35]和莫文娟等[36]的研究一致。但在花芽分化进程方面,早花类型各阶段的分化时期明显早于晚花类型,这可能是导致早花植株提前开花的原因之一。
成花基因在时间和空间上的特异表达对调控植物的开花起重要作用[10]。LFY基因是成花途径中的关键基因,从多种植物中克隆并分析了LFY同源基因的功能[37]。在拟南芥中LFY基因在营养生长阶段表达微弱,在花分生组织中有较强的表达[38]。在桃树和山核桃中同源的LFY基因主要在成花诱导期及花芽形态分化初期表达[39-40]。FT和TFL1基因均属于磷脂酰乙醇胺结合蛋白(Phosphatidyle-thanolamine-binding protein,PEBP)家族成员,具有保守的PEBP结构域,是开花调控网络下游的2个重要基因,在调节植物开花中也起重要作用。FT与TFL1功能正好相反。FT促进成花转换并启动花发育,而TFL1抑制花原基形成[41]。其原因在于FT与TFL1通过竞争性结合FLOWERING LOCUS D (FD),从而调控下游LEAFY(LFY)基因的表达[42]。FLC基因是春花途径中的关键基因,是抑制植物开花的基因,这种抑制是通过FLC基因抑制下游FT和SOC1这2个促进开花基因的表达而实现的。本研究中,PsoLFY和PsoFT基因在花芽分化前期(7月15日和8月15日),早花植株花芽中的表达高于晚花。此时,正值大多数早花植株花芽处于花芽诱导分化和花原基形成期,表明PsoLFY和PsoFT基因在新疆野生樱桃李成花诱导和花原基形成阶段起重要作用。PsoTFL基因在早花和晚花植株花芽中的表达几乎没有差别,表明该基因对开花时间的影响作用较小。而PsoFLC基因在7月15日的早花花芽中其表达量高于晚花类型,在其后的花芽分化阶段都是晚花植株中的表达高于早花植株,并且这种差异随着环境温度的降低逐渐加大。表明PsoFLC基因在环境温度较高的时期对花芽分化的影响作用较小,而在温度较低的阶段其抑制花芽分化的作用更明显。
植物在环境胁迫下通常为了逃避逆境会加速生长提前开花,使植物能够适当地缩短生长周期以应对胁迫[43]。在拟南芥中干旱胁迫可以加速植株开花,脱落酸(ABA)结合因子ABF3和ABF4通过转录调节花整合抑制因子FCONSTANS1(SOC1)基因的过度表达来调节干旱时的开花。表明ABFs和SOC1通过介导ABA信号加速开花,从而减轻干旱胁迫对繁殖产生的不利影响[44]。另外ABA还可以通过促进FLC的表达延迟开花[45]。本研究对PsoRPM2基因引起转基因植株提早开花的现象研究发现,60%的转psoRPM2基因植株表现为早花,同时转基因烟草花芽中的LFY基因表达量高于对照;而其余成花基因在转基因烟草中的表达与对照差异不大,表明在psoRPM2基因存在的植株中LFY基因可能对开花起着重要作用。同时对新疆野生樱桃李早花与晚花单株分析psoRPM2基因的表达发现,在花芽分化的诱导期(7月15日)早花花芽中的psoRPM2基因表达明显高于晚花,并且在7月15日和8月15日早花花芽中的PsoLFY和PsoFT基因表达也高于晚花,说明psoRPM2基因可能与这2个基因存在着某种联系。通过酵母双杂实验表明PsoRPM2与PsoLFY可以发生互作,但PsoRPM2与PsoFT之间没有互作(结果未列出)。表明PsoRPM2可以通过与PsoLFY发生互作影响开花。
综上,本研究表明新疆野生樱桃李早花的原因在于花芽分化诱导期通过PsoRPM2与PsoLFY互作,提高了花芽中PsoLFY和PsoFT基因的表达,加快了花芽形态分化进程,从而导致植株早花。
猜你喜欢
金山(2021年10期)2021-11-02
落叶果树(2020年3期)2020-06-17
河北果树(2020年1期)2020-02-09
热带生物学报(2019年1期)2019-04-18
计算机应用(2018年10期)2018-11-22
现代园艺(2017年23期)2018-01-18
华南师范大学学报(自然科学版)(2017年4期)2017-09-11
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
广东农业科学(2016年3期)2016-07-27
湖北农业科学(2014年6期)2014-07-02
