
磷酸盐对高水分挤压组织化植物蛋白产品品质的影响
2023-11-16 11:15安红周郭益廷李盘欣黄亚男
中国粮油学报 2023年9期
安红周, 黄 山, 郭益廷, 李盘欣, 黄亚男
(河南工业大学粮油食品学院1,郑州 450001)(小麦和玉米深加工国家工程研究中心2,郑州 450001)(河南省南街村(集团)有限公司3,临颍 462600)
植物蛋白高水分挤压(物料水质量分数≥40%)是一种植物蛋白质构重组技术,通常是指大豆蛋白、谷朊粉等植物蛋白原料,在双螺杆挤压机内部产生的高温、高压、高剪切作用下发生变性、展开,然后在冷却模具处发生蛋白质分子重排、聚集、成型的复合加工过程[1]。高水分挤压组织化蛋白产品具有类似肉类的纤维状结构,并且组织化程度高、质地均匀一致,此外,还具有氨基酸组成全面、零胆固醇、零反式脂肪酸等优点,拥有广阔的市场前景[2]。
虽然高水分组织化蛋白具有与肉类相似的纤维结构,但相关产品的品质特性,如质构特性、持水持油特性等,与真实肉类相比仍有较大差距,目前对组织蛋白品质调控的研究主要可以分为2个方面:通过改变工艺参数,原辅料配比等对组织化蛋白品质进行调控,包括挤压机螺杆转速、各区温度、水料比例等;通过添加不同的添加剂探究其对组织化蛋白品质的影响,包括添加亲水胶体、脂类物质、无机盐离子、藻类蛋白等。
磷酸盐作为一种较为常用的食品添加剂常被加入到肉制品中,以改善肉制品的保水性,乳化性、调节pH等[6]。Gadekar等[7]研究发现磷酸盐的加入可以改变pH,增加蛋白质与水的结合,进而显著改善山羊肉的多汁性,风味和整体适口性等感官特征。此外,一些研究结果表明,磷酸盐对植物蛋白及组织化蛋白的理化特性和品质有一定的影响和改善。Aidee等[8]发现三偏磷酸钠能显著改善大豆分离蛋白的乳化活性,增加其氮溶指数,提高体外消化率,主要是因为磷酸化后的大豆分离蛋白中的疏水基团暴露,有利于油水界面中蛋白质的扩散和重排。Zhang等[9]通过研究不同条件下三聚磷酸钠对大豆分离蛋白磷酸化程度的影响,当使用磷酸化的最佳条件(pH=9,时间为3 h,三聚磷酸钠质量分数为9%)时,可显著提高大豆分离蛋白的乳化性能。Peng等[10]研究发现,添加三聚磷酸钠能促进二硫键的形成,同时促进小麦蛋白聚合,从而较为显著地改善低水分小麦组织化蛋白的纤维化程度。Wang等[11]研究发现三偏磷酸钠的加入能显著提高低水分小麦组织化蛋白的持水性,降低氮溶指数和游离巯基含量。磷酸盐的加入对植物蛋白原料和植物组织化蛋白的理化特性都有较为明显的影响,然而这些研究多集中在低水分组织化蛋白领域,目前鲜有高水分挤压条件下磷酸盐与植物蛋白的相互作用,以及这些作用对高水分组织化蛋白产品的功能与品质影响的研究报道。
研究采用双螺杆挤压工艺生产高水分组织化植物蛋白,通过添加具有相同阳离子和相似阴离子的3种磷酸盐(NaH2PO4、Na2HPO4和Na3PO4)在,研究磷酸盐的种类、浓度等对组织化蛋白质构特性、持水持油、水分分布等品质的影响规律。通过微观作用力测试、二级结构变化等研究,研究磷酸盐的加入对组织化蛋白理化性质影响规律背后的作用机制,为高水分组织化植物蛋白产业化生产过程中磷酸盐这一类添加剂的加入提供参考。
1 材料与方法
1.1 材料与试剂
大豆分离蛋白(蛋白质量分数90.5%,干基);大豆蛋白粉(蛋白质量分数55.5%,干基)、谷朊粉(蛋白质量分数82.2%,干基)、小麦淀粉、DTNB、Tris、三氯乙酸、甘氨酸、EDTA、考马斯亮蓝G250、β-巯基乙醇。
1.2 仪器与设备
CLEXTRAL Ev025型双螺杆挤压,CenLee20K台式高速离心机,CR-400色彩色差仪,Kjeltec 8400全自动凯氏定氮仪,UV762紫外可见分光光度计,LGJ-10冷冻干燥机,TA-XT PLUS物性测试仪,NICOLET 6700傅里叶红外光谱仪。
1.3 实验方法
1.3.1 挤压组织化蛋白的制备
采用双螺杆挤压机[2]。原辅料按照大豆蛋白粉∶大豆分离蛋白∶谷朊粉∶小麦淀粉=4∶3∶3∶1质量比例进行混合,NaH2PO4、Na2HPO4、Na3PO4分别按添加量(质量分数,下同)0.0%、0.2%、0.4%、0.6%、0.8%、1.0%的比例加入后与原料混合均匀。
挤压参数:挤压机温度Ⅰ区30 ℃,Ⅱ区90 ℃,Ⅲ区120 ℃,Ⅳ区140 ℃,Ⅴ区150 ℃,Ⅵ区160 ℃;冷却温度60 ℃;实际喂料量2.6 kg/h;加水量3.0 L/h;螺杆转速280 r/min。挤压出的产品立即用真空包装机真空密封包装,低温保存。
1.3.2 质构特性
参考安红周等[12]的方法并略作修改。选取较为平整的挤压组织化蛋白,并用手术刀切制成2 cm×2 cm×0.5 cm的样品,使用物性测试仪采用TPA程序,选用直径为5 cm的探头(P/50),样品压缩形变量为50%,测试的测前、测中和侧后速度分别为2、1、2 mm/s,触发力5 g。平行测定8次以上,去掉异常值后取平均值。
1.3.3 色泽特性
采用色差仪对组织化蛋白的3个不同部位测定色泽,每个部位平行测定3次,记录L*、a*、b*、ΔE值,取平均值。L*表示明度系数,数值的范围为0~100,其能够反应样品的亮白程度;a*和b*分别表示红绿和黄蓝的彩度系数,数值处于60以内;ΔE表示各值与标准白板色调之间的差值程度。标准白板的色调值为L*=95.22,a*=-0.6,b*=4.15。ΔE值计算公式为:
1.3.4 剪切特性
组织化度为最大横向剪切力(垂直于组织蛋白纤维方向的力)与最大纵向剪切力(平行与组织蛋白纤维方向的力)的比值。参考Peng等[10]的方法并略作修改。实验选用挤压后冷却至室温的高水分组织蛋白产品,将其切割成2 cm×2 cm×0.5 cm的块状样品,进行剪切特性的测定(A/ECB探头),用组织化度来描述产品的剪切特性[12]。其测试条件为:将块状样品置于平台中央,测前速度和测后速度为2 mm/s,测试速度1 mm/s,剪切应变为75%,触发力5 g。平行测定6次以上,去除掉异常值后取平均值。
1.3.5 持水性、持油性测定
参考安红周等[13]的方法并略作修改。
持水性(WAC):准确称取3.00 g冻干样品倒入25 mL离心管中,加入20.00 mL去离子水,使用涡旋振荡器充分混匀,室温水平静置30 min。4 500 r/min下离心10 min,将离心管开盖后倒置于滤纸上进一步沥干多余水分,10 min后称重,所有样品均进行3组平行测试,取平均值。持水性大小以每克样品吸附水的质量表示。
式中:m2为离心管与沉淀物的合质量/g;m1为离心管与样品的合质量/g;m0为冻干样品的质量/g。
持油性(OAC):准确称取3.00 g冻干样品倒入25 mL离心管中,加入20.00 mL大豆油,使用涡旋振荡器充分混匀,室温下水平静置30 min。4 500 r/min下离心20 min,离心结束后需立即倒掉离心管中上层油脂,防止互溶,随后称质量。所有样品均进行3组平行测试,取平均值。持油性大小以每克样品吸附油的质量表示。
式中:m2为离心管与沉淀物的合质量/g;m1为离心管与样品的合质量/g;m0为冻干样品的质量/g。
1.3.6 蛋白质溶解度测定
参考陈锋亮等[14]的方法并略加修改,配置8种浸提溶液:pH=7.00的0.1 mol/L磷酸盐缓冲液(P):天然状态蛋白;P+8 mol/L尿素(P+U):破坏氢键;P+1.5 g/100 mLSDS(P+S):破坏疏水相互作用;P+0.1 mol/Lβ-巯基乙醇(P+M):破坏二硫键;P+8 mol/L尿素+1.5 g/100 mLSDS(P+U+S);P+8 mol/L尿素+0.1 mol/Lβ-巯基乙醇(P+U+M);P+1.5 g/100 mLSDS+0.1mol/Lβ-巯基乙醇(P+S+M);P+8 mol/L尿素+1.5 g/100 mLSDS+0.1 mol/Lβ-巯基乙醇(P+U+S+M)。
称取0.50 g冻干后样品放于25 mL离心管中,分别加入10.00 mL 8种不同浸提液,混匀后于振荡器水平振荡2 h,4 500 r/min离心10 min,保留上清液。使用考马斯亮蓝法测定上清液中的可溶性蛋白含量,凯氏定氮法测定产品的总蛋白含量。蛋白溶解度以上清液中蛋白含量比上总蛋白含量表示。所有样品均进行3组平行测试,取平均值。
1.3.7 游离巯基和二硫键含量的测定
参考薛腊梅[15]的方法进行测定。
试剂A(Tris-Gly-EDTA缓冲液):准确称取10.4 g三羟甲基氨基甲烷(Tris)、6.9 g甘氨酸(Gly)、1.2 g乙二胺四乙酸(EDTA)溶于1 L蒸馏水中,调节pH=8.00;试剂B(Ellman溶液):称取40 mg的2-硝基苯甲酸,用试剂A定容至10.00 mL;试剂C:β-巯基乙醇;试剂D:12%三氯乙酸游离巯基含量:准确称取3 mg蛋白样品,加5 mL试剂A缓冲液20 ℃下反应30 min,再加入40 μL试剂B,振荡混匀,25 ℃下避光静置30 min。空白为无蛋白样品,取反应后的上清液在412 nm处测得吸光度值。所有样品均进行3组平行测试,取平均值。
总巯基含量:准确称取3 mg蛋白样品,加入5 mL试剂A缓冲液,再加入0.05 mL试剂C,25 ℃下保温1 h后加入10 mL试剂D,再在25 ℃保温1 h,4 000 r/min离心10 min,倒掉上清液,保留沉淀,再加入5 mL试剂D洗涤沉淀,5 000 r/min离心10 min,再重复洗涤操作。测定沉淀中游离巯基含量即为总巯基含量。所有样品均进行3组平行测试,取平均值。

1.3.8 傅里叶变换红外光谱分析(FT-IR)
参考Qiao等[16]的方法,准确称取1 mg蛋白样品,及100 mg溴化钾,研磨混匀后,使用液压机将其制成1~2 mm厚薄片,分辨率为4 cm-1条件下共进行64次扫描,扫描范围400~4 000 cm-1,使用Omnic软件分析光谱中的峰信号,再使用pickfit软件进行拟合处理求出蛋白质二级结构含量。所有样品均进行3组平行测试,取平均值。
1.3.9 低场核磁共振测量
取70 g组织化蛋白样品,切成3 cm长条状,用聚四氟乙烯膜将其包裹放入检测管内,治愈核磁共振检测槽室进行检测。检测参数:采样点数TD=2 048,重复扫描次NS=32,弛豫衰减时间T0=10 000 ms。利用CPMG脉冲序列测定样品的自旋弛豫时间T2[17]。所有样品均进行3组平行测试,取平均值。
2 结果与分析
2.1 磷酸盐对组织化蛋白内部结构的影响
通过比较不同添加量磷酸盐处理后的样品发现,NaH2PO4和Na2HPO4相较于空白都具有较为致密的纤维结构,其中纤维结构随着NaH2PO4添加量的增大呈现先增大后减小的趋势,在添加量(质量分数,下同)为0.4%时纤维结构最为致密。当添加Na2HPO4时,也可以观察到较为明显的纤维结构,其中,在质量分数为0.2%时纤维结构最为致密均匀,而随着添加量的继续增大,内部纤维趋于片状形态,致密的丝状纤维减少。当添加Na3PO4时,产品内部纤维结构相较于空白减少,内部表面相较于空白更为光滑平整。
2.2 磷酸盐对高水分植物蛋白产品质构特性的影响
如表1分析可知,NaH2PO4和Na2HPO4对组织化蛋白的硬度无较为规律性影响,而Na3PO4的加入显著地提高组织化蛋白的硬度,呈现先增高后降低的趋势:当质量分数为0.6%时,硬度、弹性均达到最大值;当质量分数为0.8%时,咀嚼性、黏性及内聚性达到最大。这与安红周等[18]研究相一致:在碱性条件下,组织化蛋白的硬度、弹性、咀嚼性均显著增高,且随着碱性的增大呈现先升高后下降的趋势。这可能是由于在碱性条件下,会提高熔融状态下蛋白的黏度,使物料在挤压套筒内受到更大的剪切作用,后经冷却压缩阶段蛋白重新交联形成更为致密的网络结构[19]。而随着磷酸钠添加量的继续增加,在高浓度离子的作用下,蛋白质分子与离子产生相互作用增强,降低蛋白凝胶网络的交联,从而导致硬度、咀嚼性等显著下降[20]。
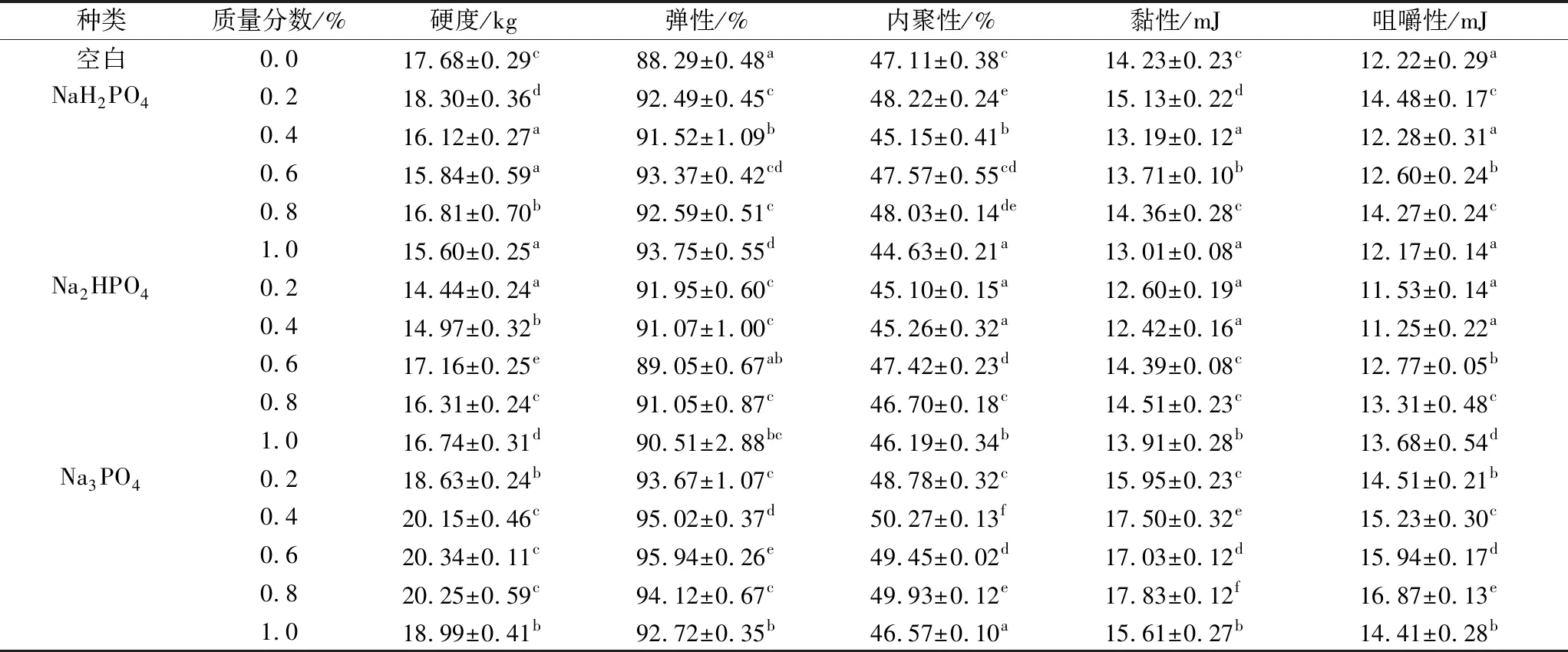
表1 磷酸盐对组织化蛋白质构特性的影响
2.3 磷酸盐对组织化蛋白色泽的影响
由表2分析可知,随着NaH2PO4和Na2HPO4的加入组织蛋白的明度系数L*显著增加,色差值ΔE显著降低,颜色较空白更为明亮。挤压组织化蛋白的颜色产生过程可能是原料中的淀粉经过水解生成还原糖,还原糖中的羰基与蛋白中的氨基在高温高压下发生美拉德褐变反应。而加入弱酸性的NaH2PO4在一定程度上会抑制美拉德反应的进行,降低美拉德反应速率,难以形成吡嗪类物质,从而导致产品明度系数L*显著增大,色差值ΔE显著降低。而当加入弱碱性的Na3PO4有利于美拉德反应的发生,在碱性条件下,氨基会水解以阴离子形式存在,此时氨基反应活性较强,较高的pH也会使还原糖中的脱羧反应增强,从而有利于美拉德反应的发生,使挤出物的明度系数L*显著降低,色差值ΔE显著升高。

表2 磷酸盐对组织化蛋白色泽的影响
2.4 磷酸盐对组织化蛋白产品组织化度的影响
组织化度通常可以用来描述产品的剪切特性,是评价组织化蛋白产品品质的重要指标之一,多用来形容挤出产品的纤维化程度。蛋白质网络结构具有一定程度上的有序性和取向性,如果蛋白质分子变性的速率比聚集的速率快,蛋白质分子就更易发生有序结合,当蛋白质分子变性速率小于聚集速率时,蛋白质就易形成无序粗糙的凝胶网络结构。而金属离子通常可以通过影响蛋白质变性的速率,从而影响蛋白质网络结构的形成,对组织蛋白纤维结构产生一定的影响[21]。组织化蛋白产品的组织化度随磷酸盐的种类和添加量的变化如图1所示。随着NaH2PO4的和Na2HPO4的加入,组织化度均显著增加,且呈现先增大后减小的变化趋势,分别在质量分数为0.2%、0.4%时达到最大值。这可能是由于在较低浓度时,磷酸盐与蛋白质分子间的相互作用,有助于蛋白质变性、打开,在挤压产生的高剪切作用下,形成致密有序的凝胶网络结构,同时在冷却分子重排时,较低的离子浓度能够促进形成更为密集有序纤维分布,显著增大挤出产品的组织化度。而随着浓度继续增大,离子强度过高时,变性的蛋白质趋向于形成随机且粗糙的纤维结构。当加入Na3PO4时,产品的组织化度呈现波动变化趋势,质量分数为0.2%时达到最大值,其原因可能是在碱性条件会增大熔融状态下物料的黏度[19],影响其在挤压套筒内的停留时间,同时较高的离子浓度会促进随机、无序结构的形成,在2种因素的共同作用下,产生了波动性的变化趋势,这与Li等[5]观察到的现象相似。
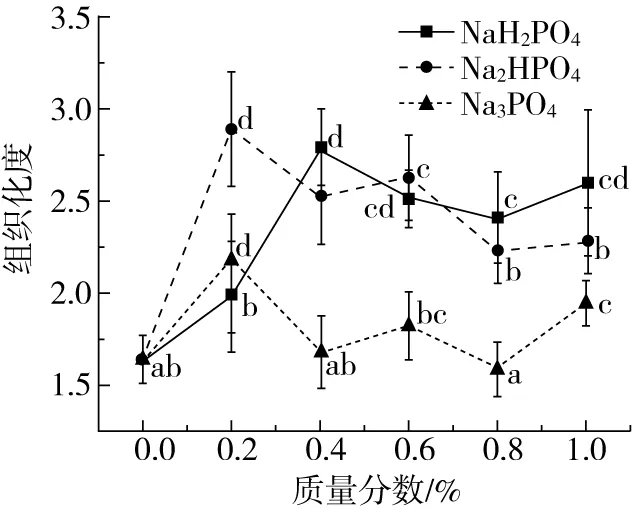
图1 磷酸盐对组织化蛋白组织化度的影响
2.5 磷酸盐对组织化蛋白持水性、持油性的影响
磷酸盐对组织化蛋白产品持水、持油特性的影响规律如图2所示。3种磷酸盐的加入,较为显著地提升组织化蛋白的持水性。其中加入NaH2PO4提升较为明显,在质量分数为0.6%的时持水性最好,随着浓度的继续升高有缓慢降低的趋势。Na3PO4的持水性,也是呈现先增高后降低的趋势。而Na2HPO4这可能跟磷酸盐可以通过在面筋蛋白和淀粉之间起到架桥结合的作用,形成稳定的复合体,增强面筋蛋白的吸水能力,从而使组织蛋白的持水性能提高[22]。而当离子浓度较高时,会破坏蛋白质的结构被破坏,从而影响蛋白质与水的相互作用,使持水性降低[23]。

图2 磷酸盐对组织化蛋白持水、持油性的影响
随着NaH2PO4的加入,组织化蛋白的持油性呈现先升高后下降的趋势,在质量分数为0.4%时达到最好的持油性。NaH2PO4和Na3PO4的加入都显著降低了组织化蛋白的持油性。可能原因是磷酸盐的加入,促进了引起蛋白质网络结构变化,减少了疏水位点与油结合,导致持油性下降[24]。总体来看,NaH2PO4和Na2HPO4的加入可以优化组织化蛋白的微观结构,提升产品整体的持水持油特性。
2.6 组织化蛋白溶解度分析
蛋白质在不同浸提液中的溶解度可以反映蛋白质分子的变性程度,以及维持蛋白质微观结构的主要作用力,蛋白质溶解度越高,表明该浸提液所破坏的化学键或者微观作用力在维持蛋白结构中起的作用越强[25]。组织化蛋白产品在浸提液中不同的溶解度表明了维持蛋白质分子结构的微观作用力具有多样性。
磷酸盐的添加对组织化蛋白溶解度的影响如图3所示。单一作用破坏试剂(P+U、P+S、P+M)实验表明,氢键是维持组织蛋白微观结构的主要作用力,并且磷酸盐的加入并未对氢键造成的溶解性变化产生显著性影响;二硫键和疏水相互作用等化学键或微观作用对蛋白质溶解性几乎没有影响,这与Wei等[26]的研究结论相似。而在氢键、二硫键和疏水相互作用共同作用下的蛋白质溶解度变化为负值,这是因为尿素溶液不仅会破坏氢键,还会对疏水相互作用有一定程度的干扰,导致重复计算氢键和疏水相互作用。此外当氢键和二硫键的协同作用导致蛋白质溶解度显著增大,可能是由于在构成蛋白质结构过程中,氢键是维持蛋白质分子的基础作用力,且氢键的排列更为有序紧密,保护了位于结构内部的二硫键交联结构,当浸提液中的尿素将氢键破坏后,内部的二硫键也随即被巯基乙醇破坏,导致蛋白质溶解度显著增加。
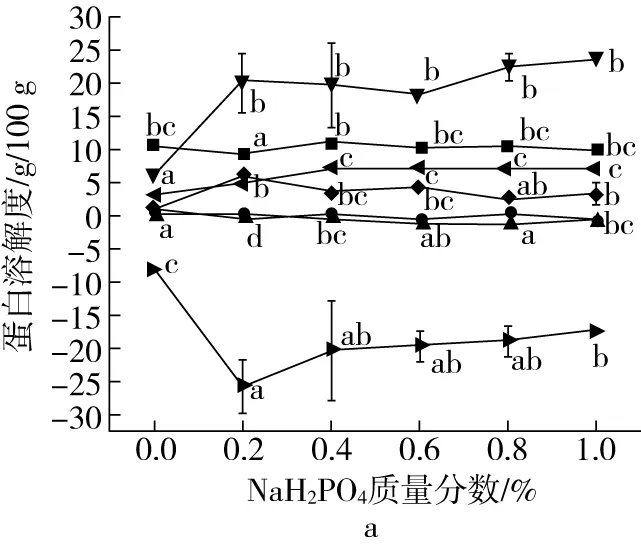
图3 磷酸盐对组织化蛋白溶解度的影响
2.7 磷酸盐对组织化蛋白产品中水分分布的影响
低场核磁广泛的被用于表征食物中水的状态和分布情况。T2是激发态的自旋-自旋质子与相邻质子交换能量以达到动态平衡所花费的时间,又称弛豫时间。较长的T2弛豫时间表示水分子具有更大的自由度,而较短的T2弛豫时间意味着较低的自由度。根据T2弛豫时间曲线出峰的位置,通常将水的状态分为3种:T2b(0~10 ms)、T21(1~100 ms)和T22(100~300 ms),分别对应着强结合水(强结合质子),弱结合水(中结合质子)和自由水(弱结合质子)[33]。A2b、A21和A22为3种状态水对应的峰面积,峰面积的大小表示该状态水含量的多少。
图4为组织化蛋白产品中水分子的弛豫曲线(以0.6%为例),在0~10 ms内出现了2个T2b峰,表明有2种结合程度不同的强结合水,这可能是因为原料中同时含有蛋白质和淀粉。强结合水通常通过氢键、静电等强相互作用与食品大分子中的羧基、氨基或者羟基结合。10~100 ms出现的T21峰为主峰(含量比例超过80%),这部分水为弱结合水,与强结合水相比流动性受限,通常是指被蛋白质网络结构包裹着的水[34]。样品中水分主要是以弱结合水的形式存在的,表明了组织蛋白具有良好的持水性。100~300 ms出现的T22峰表示为自由水,表示可以在产品组织内可以自由流动的水。

图4 磷酸盐对组织化蛋白水分分布的影响
由图4和表3可知,组织化蛋白中自由水含量占比很低,说明组织蛋白内的自由水含量非常少,大部分都是以结合水的形式存在,不利于产品形成多汁的口感。同时由表3可知,添加3种磷酸盐都增加强结合水含量,这可能是磷酸盐中的无机盐离子的离子作用和水合作用,吸附了部分水分子,导致强结合水含量增大[35]。

表3 磷酸盐对组织化蛋白水分分布的影响
2.8 组织化蛋白分子中游离巯基和二硫键含量分析
组织化蛋白分子中游离巯基和二硫键的变化如图5所示,结果表明,磷酸盐的加入能够促使游离巯基转化为二硫键,增强蛋白质分子的交联。3种磷酸盐的添加后,游离巯基含量较空白均显著降低,二硫键含量显著升高,但均呈现出先增加后降低的趋势。这可能是金属离子的静电作用和酸碱特性共同影响下的结果。Zhang等[27]发现高浓度的盐加入,水分子优先跟金属离子结合,使得蛋白质周围水分子重新排列,蛋白质分子的疏水相互作用增强。金属离子的加入影响蛋白之间的微观相互作用,使物料黏性增大,在挤压套筒内停留时间增长,变性更为彻底,从而促进了游离巯基向二硫键的转换。而Na2HPO4和Na3PO4的添加导致的游离巯基和二硫键的波动变化,原因可能是碱性条件会促进巯基的氧化反应,诱导游离巯基向二硫键发生转换[28],而随着碱性的增加,游离巯基含量呈现缓慢上升趋势,这可能是由于,在碱性条件下胱氨酸发生了β-消去反应,生成了脱氢丙氨酸和巯基,从而使游离巯基得到了补充[29]。
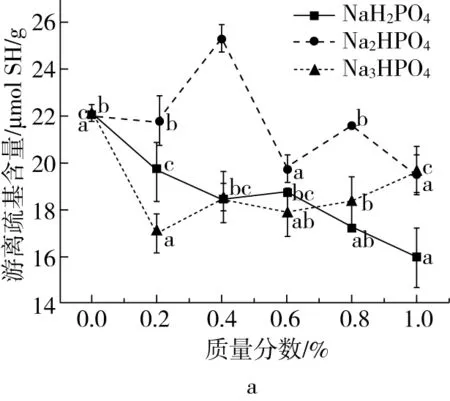
图5 磷酸盐对组织化蛋白中游离巯基、二硫键的影响
2.9 傅里叶红外变换光谱分析
酰胺Ⅰ带(1 600~1 700 cm-1)常用来描述蛋白质结构,并广泛用于定量分析蛋白质的二级结构,主要是由肽链上的C—O伸缩振动引起的[30]。在酰胺Ⅰ带1 650~1 660、1 610~1 640、1 660~1 700、1 640~1 650 cm-1这4个波长处的吸收峰分别对应蛋白质分子结构中的α-螺旋、β-折叠、β-转角和无规则卷曲[31]。由表4可知,不同磷酸盐的加入对组织化蛋白的α-螺旋和β-转角均无显著的规律性影响,而加入NaH2PO4能提升β-折叠结构且降低无规则卷曲含量,说明NaH2PO4的加入使部分无规则卷曲转变为β-折叠结构,产品结构更加趋于有序。β-折叠结构的增加是由于分子间反平行β-折叠的形成,而反平行β-折叠结构常见于蛋白质的热变性聚集过程中,这也印证了NaH2PO4的加入有利于产品组织化度的增大[32]。而随着Na2HPO4的加入,β-折叠含量呈现先增大后减小的趋势,与之对应的无规则卷曲呈现先减小后增大的趋势。而随着Na3PO4的加入,对组织化蛋白β-折叠和无规则卷曲无显著规律性影响。这说明在挤压过程中存在β-折叠和无规则卷曲的相互转化,NaH2PO4和Na2HPO4的加入有利于产品β-折叠结构含量的增加和无规则卷曲的减少,进而对产品组织化度、质构特性等产生影响。
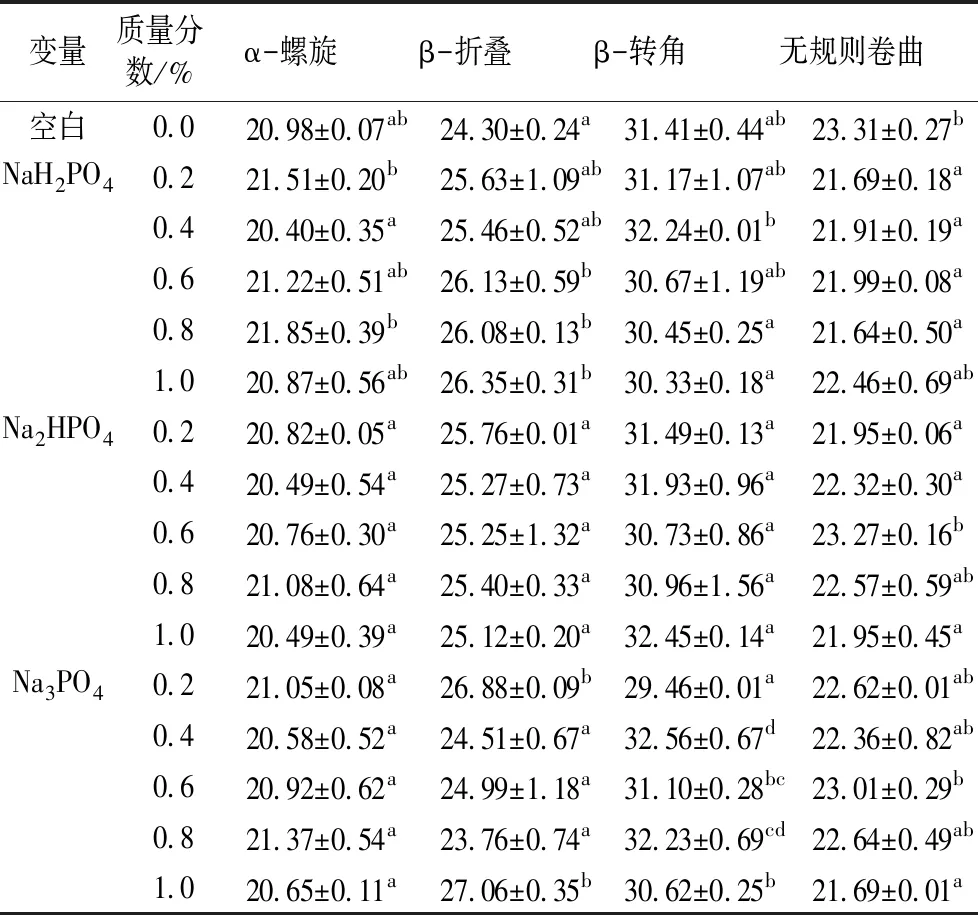
表4 磷酸盐对组织化蛋白分子二级结构的影响
3 结论
通过控制NaH2PO4、Na2HPO4和Na3PO4的添加量,研究3种磷酸盐的加入对组织化蛋白理化性质影响,发现:添加NaH2PO4和Na2HPO4在质量分数为0.2%~1%内都显著提升产品的组织化度,Na3PO4的加入显著增大了产品的硬度、弹性和咀嚼度。3种磷酸盐的加入都显著改善了组织化蛋白的持水性。磷酸盐的加入降低了产品的游离巯基含量,增大了二硫键含量,说明磷酸盐有利于游离巯基向二硫键的转化。傅里叶红外光谱分析表明,NaH2PO4和Na2HPO4的加入有利于产品蛋白中的无规则卷曲向β-折叠转换,形成更有序的结构,这也与组织化度变化相一致。蛋白质溶解性测试结果表明氢键对维持组织化蛋白结构起着重要作用,其次是二硫键,最后是疏水相互作用。低场核磁结果表明组织化蛋白中水的存在方式为弱结合水,水分与蛋白结合较为紧密,不易流失,磷酸盐的加入能够提升强结合水的含量,增加组织化蛋白产品的持水性能。磷酸盐的加入显著性的提升了组织化蛋白的持水性、剪切特性,并通过巯基二硫键、傅里叶红外光谱、低场核磁等实验结果从微观层面对其改善原因进行进一步的研究,初步揭示了磷酸盐对产品品质的影响机理。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
质谱学报(2021年6期)2021-12-06
中国生物制品学杂志(2021年10期)2021-10-20
河南工业大学学报(自然科学版)(2021年3期)2021-07-13
农业工程学报(2019年1期)2019-01-18
中国粮油学报(2018年1期)2018-03-10
中国饲料(2016年17期)2016-12-01
天津科技大学学报(2016年1期)2016-02-28
四川师范大学学报(自然科学版)(2015年1期)2015-02-28
海洋科学进展(2015年1期)2015-02-27
