
四川黑茶渥堆发酵不同时期细菌群落结构与多样性
2023-12-21 15:35严宽李翔宇张建王玉洁刘思岑ManzarAbbas李俊强
宜宾学院学报 2023年12期
严宽,李翔宇,张建,王玉洁,刘思岑,Manzar Abbas,李俊强
(宜宾学院 农林与食品工程学部,四川宜宾 644000)
黑茶是我国六大基本茶类之一,加工原料一般较为粗老.其初加工主要有4 道工序,其中渥堆是最重要的一步.渥堆进行的好坏直接决定了黑茶最终的品质,在这一过程中,由于微生物的参与,它们在其中进行代谢活动产生了很多特殊物质,这些物质造就了黑茶独特的色、香、味[1].因云南、湖南和四川等地的黑茶原料不同及生产标准的差异性,造成微生物群落在黑茶生产过程中所产生的特征风味也有所不同.周红杰[2]发现:黑茶渥堆有许多的微生物的参与,以黑曲霉最多,酵母次之.研究者发现在六堡茶的茶堆中存在着大量的细菌、酵母、霉菌;而雅安藏茶的渥堆中的真菌种群主要是黑曲霉、塔宾曲霉、总状枝毛霉、炭黑曲霉、根霉等;在茯砖茶渥堆过程中普遍存在的微生物主要是假单胞菌和克雷伯氏菌[3-5].茶叶的品质受微生物的影响非常大,在发酵的不同的阶段中将其中的微生物的配比调整为最适比例,将会极大地缩减发酵所需的时间.如果将黑茶的发酵过程的微生物种类、温度和酸碱度加以控制和规范,可以使黑茶有稳定的品质[6].另外,由于有害杂菌(如曲霉属、青霉属等[7])对黑茶品质造成影响,在黑茶的生产过程中不但要注意有益菌的生长代谢活动,同时也要时刻关注有害菌的生长活动,及时排除其对茶品质的威胁.加强对黑茶发酵中的有益菌以及有害杂菌的研究,将对黑茶的生产、品质的把控以及风味的形成至关重要.
四川作为中国黑茶的主要产区,所生产的黑茶产品因其原料生长区域有着特殊的气候条件,且在渥堆过程中的特殊环境条件和黑茶加工技术的差异,可能存在着与其它地区的黑茶完全不同的潜在微生物资源.目前对四川黑茶的研究主要针对其活性成分功能等的研究,鲜有对其渥堆发酵过程中有关微生物的报道.因此,本研究利用高通量测序研究黑茶渥堆过程的整个细菌群落结构多样性研究黑茶堆制过程中细菌群落的结构和多样性,分析其中细菌的种类和作用机制,为改进黑茶加工工艺和提高黑茶质量提供科学依据.
1 材料与方法
1.1 实验材料
黑茶样品是由四川省茶业集团股份有限公司提供夏秋季采摘的茶树鲜叶.在发酵过程中,将茶树的叶子彻底混合以确保均质,并以适量的自来水喷洒以保持65%~75%(w/v)的固体含量和45~71°C的温度.在发酵过程中重复三次并在以下时间间隔收集样品:第0(YC)、8(W1)、16(W2)、24(W3)和32(W4)天.每天记录每个发酵茶堆中心40 cm 深度的温度情况.
1.2 DNA提取
将5 g 黑茶样品在无菌条件下悬浮在50 mL Tween-NaCl 缓冲液[0.9% (w/v) NaCl、0.05% (v/v)Tween 20、2% (w/v)聚乙烯聚吡咯烷酮]中,将匀浆在4 ℃的超声仪中放置30 min.悬浮液过无菌纱布,4 ℃条件下,2000 r/min 离心2 min,去除颗粒物和上清液,在4 ℃ 12000 r/min 离心10 min,得到微生物沉淀.用E.Z.N.A.TM HP Plant DNA Kit(Omega Bio-Tek Inc., GA, USA)从沉淀物中提取微生物基因组DNA,用E.Z.N.A.TM Soil DNA Kit进一步纯化DNA制剂以去除可能的PCR抑制剂.
1.3 PCR扩增和克隆文库的建立
利用引物338F 和806R(5'-ACTCCTACGGG AGGCAGCAG-3' , 5'-GGACTACHVGGGTWTC TAAT-3')进行扩增,获得细菌16S rRNA 基因的V3+V4 区域[8-9].PCR 反应条件为:95 °C 3 min;95 °C 30 s,55 °C 30 s,72 °C 45 s,30 个循环;72 °C 10 min;10 °C 10 min.PCR 反应体系为:加入5×PCR buffer (with Mg2+) 4 μL、2.5 mmol/L 的dNTP 2 μL、0.8 μL 的5 μmol/L P1 (338F)及0.8 μL 的5 μmol/L P2 (806R),再加入0.4 μL 5 U/μL的Taq酶、2 μL DNA 模板和10 μL ddH2O.将PCR 产物混合后用2%琼脂糖凝胶电泳检测,再通过使用AxyPrep-DNA 凝胶回收试剂盒(AXYGEN 公司)切胶来回收PCR 产物,并用Tris_HCl 洗脱;再用2%琼脂糖电泳检测,对PCR 产物进行文库构建和评估,再利用Ilumina Miseq PE300 测序.根据PE reads 之间的overlap 关系,将reads 拼接(merge)成一条序列,同时对reads 的质量和拼接的效果进行检测,根据序列首尾两端的barcode 和引物序列区分样品得到有效序列,对序列方向进行校正[10].
1.4 数据处理
用Uparse OTU 聚类软件工具在97%的身份阈值下进行系统进化聚类分析,以识别具有代表性的操作分类单元序列(OTUs)[11].采用RDP classifier 贝叶斯算法对97%识别阈值下的OTU 进行分类学分析.在界、门、纲、目、科、属和种水平上创建每个样本的群落组成和科学分类[12].稀释曲线是以个体数与物种数来构建曲线,需从样本中随机抽取一定数量的个体,统计这些个体所代表的物种数目,并利用Mothur 做稀释曲线分析,利用R 语言工具制作曲线图[13].使用Chao 1、ACE、Shannon 和Simpson 指数计算每个样品的α-多样性,以评估测序深度和覆盖率,并比较不同微生物群落的丰度和多样性[14].统计多个样本中所共有和独有的OTU数目,构建韦恩(Venn)图[15];根据分类学分析结果,对单个或多个样本在各分类水平上进行比对,并使用R 语言工具构建群落结构图[16-17].
采用标准差(SD)的均值对所有数据进行解释,并采用单因素方差分析(ANOVA)进行分析.在p<0.05 显著性水平下,采用Duncan 多重比较检验检测所有样本均数之间的变异.所有相关和路径系数分析均使用和Excel 2019进行.
2 结果与分析
2.1 测序数据分析
表1 为5 个组的测序统计数据.YC-W4 五个时期的有效序列分别为62456 条、46909 条、40399条、43362 条、60400 条;检测总碱基数分别为26118006 bp、19638346 bp、17285142 bp、18361869 bp、25535185 bp;序列平均长度分别为418.59 bp、420.24 bp、427.75 bp、423.51 bp和422.57 bp.

表1 5个组样品的测序统计数据
稀释曲线(Rarefaction curve)反映了样品取样的深度.从图1 可知,这5 个组的曲线基本趋于平缓,说明取样基本合理.四川黑茶内生细菌群落结构的置信度较高,能比较真实地反映黑茶样本的内生细菌群落.

图1 所有样品的稀释曲线
2.2 OTU聚类分析
对97%相似水平下的OTU 进行生物信息统计分析,并进一步绘制Venn图(图2),以直观反映样本在不同分类水平上的组成相似性和重叠情况.对有效数据进行聚类总共得到2 948 个OTU,分为42 个门、98 个纲、247 个目、461 个科、1 052 个属和1 888个种.各个时期的样本OTU 分别为2 082、2 116、381、399 和580 个.其中,YC 与W1 阶段共有的OTU 为1 211 个,YC 与W2 阶段共有的OTU 为15个,YC 与W3 阶段共有的OTU 为25 个,YC 与W4阶段共有的OTU 为50 个.各个时期的样本独有的OTU 分别为360、399、70、94、151 个,分别占各自总数的17.3%、18.9%、18.4%、23.6%和26.0%.而5 个时期样品共有的OTU 数量为103 个,占所有样本OTU 总数的3.6%,说明在进行渥堆发酵后,四川黑茶的细菌群落结构发生了显著的变化.

图2 OTU的Venn图分析
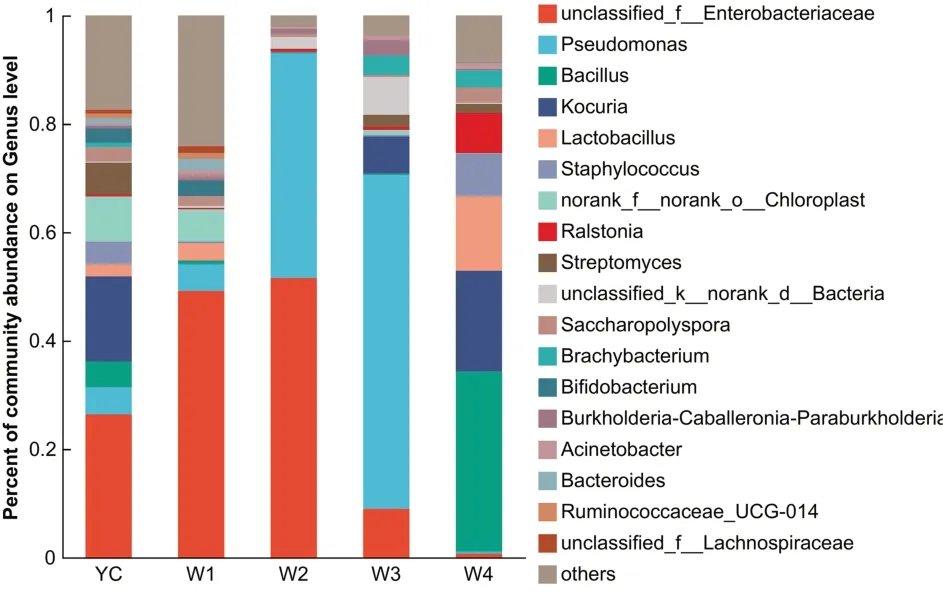
图3 属水平下的细菌群落结构柱状图
2.3 Alpha多样性分析
五个样本测序深度均在0.9984 以上,能较好地代表不同渥堆发酵时期的黑茶样本中细菌群落的真实情况(表2);一般Shannon 指数越高,Simpson 指数越低,说明样本中细菌群落多样性指数越高[18].YC和W4 两个时期的样品细菌群落多样性高,W1 和W2 时期的次之,W2 时期样品的细菌群落多样性最低.Chao 指数和ACE 指数反映了样本细菌群落的丰富度.YC时期样品的细菌群落丰富度最高,其次为W4 和W1 时期样品,W2 和W3 时期样品的细菌群落丰富度最低,这说明不同渥堆发酵时期四川黑茶样品的细菌丰富度和多样性有较大差异.
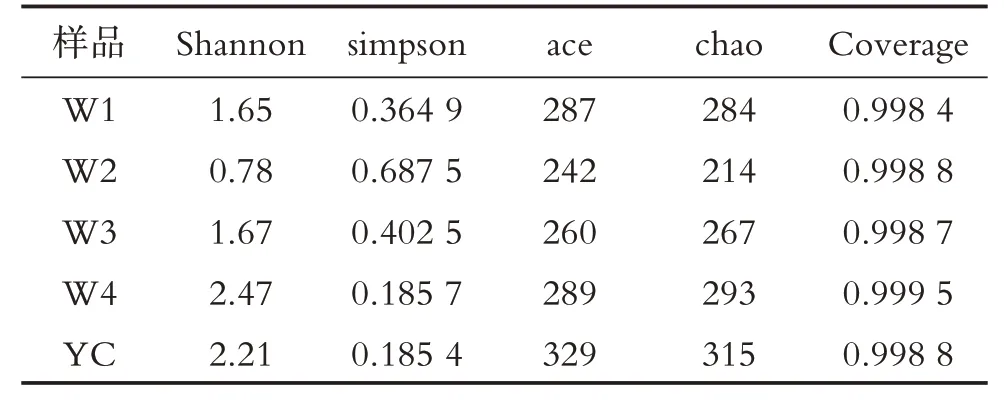
表2 四川黑茶真菌群落丰富度和多样性指数
2.4 不同时期黑茶渥堆细菌群落结构分析
在属水平上注释到明确分类地位的有18 个属,其他相对丰度较低的合并为一类(others),其中相对丰度较高的为假单胞菌、未分类肠杆菌(unclassified_f_Enterobacteriacea)、芽孢杆菌、考克氏菌属(Kocuria)、青枯菌属(Ralstonia)、乳酸杆菌、葡萄球菌、未分级叶绿体菌(norank_f_norank_o_Chloroplast)等.YC 时期,优势菌为未分类肠杆菌(26.38%),相对丰度较高的为考克氏菌属(15.72%),未分级叶绿体菌(8.32%),链霉菌(5.92%)、假单胞菌(5.02%)、芽孢杆菌(4.73%)、葡萄球菌(4.19%)等.W1时期的优势菌为未分类肠杆菌(26.38%),相对丰度较高的有未分级叶绿体菌(5.93%),假单胞菌(4.91%)、乳杆菌(3.18%)、双歧杆菌(2.90%)、拟杆菌(2.10%)等.W2 时期中相对丰度最高的仍然是未分类肠杆菌(51.55%),其次是假单胞菌(41.50%),未分类细菌(unclassified_k_norank_d_Bacteria)(2.15%),其他类细菌属相对丰度均低于1%.W3 时期中假单胞菌(61.61%)为绝对优势菌,相对丰度较高的有未分类肠杆菌(8.98%),考克氏菌属(6.86%),未分类细菌(6.97%)等.W4 时期中的芽孢杆菌(33.20%)相对丰度最高,其次是考克氏菌属(18.63%),乳杆菌(13.74%)、葡萄球菌(7.92%),青枯菌属(7.39%)等.可以发现,假单胞菌属和未分类肠杆菌在YC、W1、W2 和W4 时期中均有较高的丰度,而W4 时期主要以芽孢杆菌、考克氏菌属和葡萄球菌相对丰度较高(图4),这说明四川黑茶中细菌属的丰度随不同渥堆发酵时期的不同而存在明显差异.

图4 属水平下的群落结构热图
2.5 黑茶渥堆细菌群落组成对活性成分的影响
RDA 分析即冗余分析,是环境因子约束化的PCA 分析,将样本和环境因子反映在同一个二维排序图上,可以用来反映菌群与环境因子之间关系.从图5 可以看出,在四川黑茶的4 个活性成分中,多糖(PO)与总黄酮(FL)呈正相关关系,而咖啡因(CA)与氨基酸(AA)均呈正相关关系.进一步对黑茶活性成分、细菌分类单元和不同渥堆时期之间的相关性分析表明,咖啡因、氨基酸的有效形成主要在YC、W1、W2 时期,且与未分级叶绿体菌和未分类肠杆菌有不同程度的正相关;多糖和总黄酮的含量增加主要在W4 时期,其中芽孢杆菌、青枯菌属、糖多孢菌与总黄酮的形成相关性较高,而考克氏菌属与多糖的形成密切相关;假单胞菌是W3 时期的优势菌,但与以上四种黑茶的活性成分没有相关关系,这表明渥堆细菌群落组成在一定程度上影响着四川黑茶活性成分的合成.
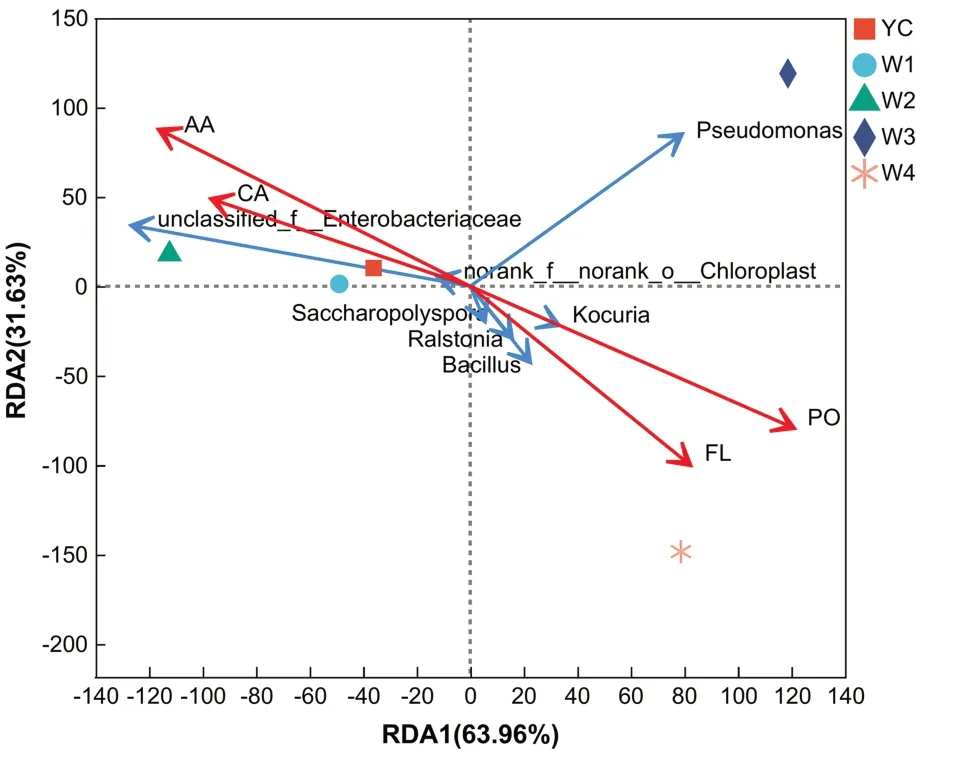
图5 属水平下的细菌RDA分析
3 讨论
四川黑茶在渥堆过程中发生了一系列复杂的变化,其渥堆周期的长短和风味特征的形成主要取决于微生物群落结构的组成及变化.传统的分离培养的方法,不仅耗时费力,而且有很多微生物无法培养出来.本实验共得到高质量细菌序列253 526条,平均长度为422.53 bp.聚类后总共得到2 948 个OTU,分为42 个门、98 个纲、247 个目、461 个科、1 052 个属、1 888 个种.这些数据表明渥堆中的活性细菌来自与渥堆相关的环境中,并且在整个发酵过程中将环境细菌接种到了茶堆中.因此本研究采用高通量测序技术探究了四川黑茶渥堆过程中细菌群落结构的组成及变化特征,有助于准确反映微生物种群数量.
近年来,对于黑茶中微生物群落组成的研究主要集中于以冠突散囊菌等为主的真菌,而对于黑茶中细菌群落组成的研究较少.Liu[19]等发现黑茶中细菌种类较为丰富.在发花早期,由于冠突散囊菌的生长,细菌的生长受到一定抑制,而在发花后期细菌总数量呈上升趋势,与冠突散囊菌保持共生关系.Zhao[20]等发现不同地域黑茶细菌群落多样性存在一定的差异.本实验通过对四川黑茶渥堆中细菌群落多样性的研究,获得了大量的细菌资源.结果表明,在属的水平上,由YC 到W3时期的样品中检测到了2 个相对丰度较高的属,分别是未分类肠杆菌属和假单胞菌,其中未分类肠杆菌属在W2 时期样品中的相对丰度最高,达51.55%.肠杆菌属在自然界中分布广泛(例如在土壤、水、植物、昆虫和动物中),肠杆菌属在黑茶渥堆发酵的前期占主导地位,且大多是无害的.黑茶中含有的假单胞菌、弧菌、葡萄球菌等通常被认为属于条件致病菌[21],其中假单胞菌对人体有一定的坏处,如通过感染伤口进入人体后,通过血液循环导致人类得菌血症.在渥堆中产生或者某个时期生长比较旺盛的有害菌,他们在随着渥堆的不断进行导致茶堆的环境发生变化,抑制其的生长,能够在一定程度上保证茶叶品质.假单胞菌在W3 时期样品中的相对丰度最高,为61.61%.且在各个时期的黑茶样品中均有较高丰度,为确定它们是否是病原体,其在黑茶中的作用和安全性还有待进一步研究.
本研究发现W4 时期的细菌群落多样性程度较高,是四川黑茶品质形成的关键时期.W3 时期的样品中芽孢杆菌是优势菌,丰度达到33.20%.这可能是因为温度逐渐降低,利于其生长.结果表明,在发酵的最后阶段,温度降低,细菌多样性增加,细菌群落的组成可能与发酵温度有关,这表明我们可以通过控制黑茶渥堆的温度来控制微生物的分布.例如,通过在最后阶段保持50℃,然后对茶砖进行快速干燥,可以将芽孢杆菌的丰度保持在高水平.有趣的是,芽孢杆菌属中有许多是益生菌,对动物和人类的肠道环境,粪便频率和特征以及皮肤特性均显示出有益的作用[22].其中凝结芽孢杆菌(Bacillus.coagulans)已作为一种食品成分进行了安全性评估,并被欧洲食品安全局添加到其合格安全性推定(QPS)清单中,并获得批准美国食品药品监督管理局公认的安全标准(GRAS).由于凝结芽孢杆菌是黑茶发酵过程中的主要细菌,又是一种安全的益生菌,可以用以改善某些食品的风味和保质期.
本研究发现四川黑茶中部分活性成分如多糖、黄酮、咖啡因、氨基酸的含量和一些细菌属丰度的相关性较高.在渥堆的YC、W1、W2 阶段氨基酸和咖啡因的含量大大增加,未分类肠杆菌和未分级叶绿体菌是这一时期主要菌种,它们与氨基酸和咖啡因的形成有关,W3 阶段假单胞菌是其中的优势菌,在W4 阶段多糖和总黄酮的含量增加,其中总黄酮与芽孢杆菌、青枯菌属、糖多孢菌的形成相关性较高,而多糖与考克氏菌属的形成密切相关.有研究表明,黑茶的活性成分的产生与细菌的代谢有着密不可分的关系,渥堆过程中多糖、总黄酮、氨基酸等活性成分存在一定程度的下降,这是由于微生物代谢的结果,普洱茶中也存在类似的现象[23].微生物代谢所分泌的纤维素酶、半纤维素酶、蛋白酶等对主要代谢产物具有催化作用[24].在后续的研究中还需要进一步探究黑茶的活性成分以及微生物的多样性,明确微生物在渥堆过程中的代谢机制,为日后对渥堆不同阶段的调控提供依据,最终提升黑茶生产工艺和品质.
4 结语
四川黑茶因其独特的香气而在经济上占有重要地位.微生物在其独特的香气、营养价值和品质的发展中起到至关重要的作用.对渥堆阶段特定细菌组成的深入了解将为精确接种益生菌以进行生物强化提供标准.细菌16S rRNA 基因的高通量测序显示,未分类肠杆菌的某些成员在YC、W1和W2早期渥堆阶段占优势;假单胞菌在W3 阶段占优势;最高的细菌多样性在W4 阶段.我们观察到益生菌菌属,如芽孢杆菌属、乳杆菌属、双歧杆菌属和糖多孢菌属在W4 的最后渥堆阶段大量存在.总之,在黑茶中有效接种这些细菌属的成员可能会提高其营养价值.
猜你喜欢
华中农业大学学报(自然科学版)(2022年5期)2022-10-13
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
茶叶通讯(2019年3期)2019-03-27
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
中国茶叶加工(2015年6期)2015-11-20
水生生物学报(2015年1期)2015-02-28
现代检验医学杂志(2015年1期)2015-02-06
食品工业科技(2014年23期)2014-03-11
