
油樟茎部内生真菌多样性分析及功能筛选
2023-12-21 15:35刘雯雯胡连清牟嘉勇蒙昌廷张淑瑶舒豪冯瑞章魏琴周万海
宜宾学院学报 2023年12期
刘雯雯,胡连清,牟嘉勇,蒙昌廷,张淑瑶,舒豪,冯瑞章,魏琴,周万海
(1.宜宾学院 四川省油樟工程技术研究中心,四川宜宾 644000;2.宜宾学院 农林与食品工程学部,四川宜宾 644000)
植物内生真菌是指长期或生活史某一个阶段定殖于健康植物内部而不引起宿主明显病理症状的微生物,利用组织学方法可从表面严格消毒植物组织中分离纯化出来[1-2].研究表明,内生真菌种类和数量会因不同宿主或同一宿主的不同地域、季节或器官等而不同[3],同时内生真菌与宿主长期共存条件下已建立互利共生关系,促进植物生长、增强宿主抗病性和环境适应的能力.部分内生菌可以产生与宿主相同或相似的活性物质,对植物的种群结构、健康状态和生存具有重大影响,现已证实它在农业、医药、发酵工程、生物防治等方面具有较大潜力[4].近年来,已对丹参[5]、芦苇[6]、黄芪[7]等不同生境植物内生真菌展开了研究.内生真菌可作为新的资源来替代短缺的药用植物并保护濒临灭绝的天然植物物种[8],故研究内生真菌多样性对了解其与宿主的生态演化、发掘活性功能菌株等具有重要意义.
油樟(Cinnamomumlongepaniculatum)属樟科(Lauraceae)樟属(Cinnamomum),是我国特有的香料林业资源,主要分布于四川省宜宾市,面积达3.13×104hm2,已成为当地主导产业之一.油樟根、茎、叶中富含松油烯4-醇、1,8-桉叶油素和α-松油醇等重要化学成分,可用于香精香料、食品添加剂、医药等行业,具有重要经济价值.关于油樟内生菌的多样性和功能性已有许多研究,如冯瑞章等[4]从油樟中筛选到3 株具有溶磷作用的内生细菌,对植物的生长、抗性和生物防治具有应用效果;严宽等[9]的研究显示利用油樟内生真菌孢子悬浮液能显著提高叶片的挥发油含量;谭韵雅等[10]发现YY26、YG42油樟内生真菌可促进1,8-桉叶油素等成分的积累物质,但会抑制YG71 菌株中代谢产物的分泌.目前内生菌的研究主要集中在油樟叶和根部位,而未见关于不同季节油樟茎内生真菌的分布情况与多样性的分析,基于此,本研究以四川宜宾地区不同生长季节油樟茎为材料,分析茎部内生真菌的分布情况与多样性,并对分离纯化菌株进行产黄酮、抗氧化、产酶等功能挖掘,以期为油樟内生真菌功能的深度挖掘、内生真菌资源多样性及提供可应用的功能菌株奠定基础,促进更有效地保护和开发油樟资源,从而缓解油樟资源匮乏和生态环境恶化.
1 材料与方法
1.1 材料
油樟采自四川省宜宾市高县(28°47'77″N、104°59'76″E)油樟优良品系谱,根据前期不同株系产樟油量的评价,选取5 株产油性能差异大的株系为材料,分别于春、夏、秋、冬各季于此5 株油樟树体上、中、下部随机采摘足够的茎,混合均匀后,用冰盒立刻带回实验室,存于4℃冰箱.马铃薯葡萄糖琼脂培养基(PDA)、麦芽提取物琼脂培养基(MEA)、卵磷脂吐温80 营养琼脂培养基用于内生真菌分离纯化.
1.2 方法
(1)油樟茎部内生真菌的分离纯化.将不同季节油樟茎分别进行消毒,消毒步骤为:无菌水冲洗3次,其次用95%酒精浸泡1 min,再用4%次氯酸钠浸泡5 min 后,用95%酒精浸泡1 min,无菌水冲洗4 ~6 次,最后用无菌剪刀将茎剪成2 ~ 3 mm 小段.取最后一次冲洗液0.1 mL 涂布于PDA 培养基平板上,26℃恒温箱中培养5 d,检测表面消毒效果.将剪好的茎段接种到培养基中,需均匀地接种5 个茎段.置于恒温培养箱中26℃ 5d 培养,观察空白培养基,若未出现菌落,则开始进行纯化.根据不同菌株生长周期的不同,分别一段时间内先后进行3 次挑菌.通过形态学分析法,挑选形态、颜色、大小不同的菌落用三点法(一个平板上等距离接种三个地方,大致呈三角形状)挑至对应的培养基上,直至得到三个纯的单菌落的平板,封好置于4℃冰箱保存备用.
(2)内生真菌的ITS 序列分析.利用真菌DNA提取试剂盒提取纯化菌株的DNA,以真菌通用引物ITS1(5'-TCCGTAGGTGAACCTGCGG-3')和ITS4(5'-TCCTCCGCTTATTGATATGC-3')为引物,扩增ITS 保守区域.PCR 反应体系包括: 2 μL DNA模板、1 μL ITS1(10 mM)、1 μL ITS4(10 mM)、25 μL 2×Taq PCR MasterMix、21 μL ddH2O.PCR 扩增程序为:94℃预变性5 min,94℃变性35 s,58℃退火55 s,72℃延伸45 s,共45 个循环,72℃再延伸10 min.PCR 产物用10 g/L 琼脂糖凝胶电泳进行检测.然后送检公司为上海生工生物工程技术服务有限公司测序.将所得序列信息提交至NCBI 网站BLAST程序进行序列同源性比对.
(3)油樟茎内生真菌多样性分析.选用4 个指标评价油樟茎内生真菌的多样性,即物种丰富度S、Shannon-Weiner 多样性H'、Simpson 优势度D、均匀度E,计算公式为:
式中:N为样本全部属数之和,Pi为属相对频率,指样本中分离某属的菌株数占分离总菌株数的百分比.
(4)内生真菌产黄酮菌株的筛选和形态学鉴定.参考柯树炜等[11]方法对菌株发酵液和菌丝体中次生代谢产物进行提取,4℃保存备用.选择氯化铝溶液、氢氧化钠溶液、氯化铁溶液和盐酸-镁粉溶液[12]显色反应初步筛选产黄酮菌株,并利用紫外分光光度法[13]测定黄酮产量.
(5)产黄酮菌株的其他功能分析.采用福林酚法测定菌株菌丝体和发酵液中多酚的含量[14].参考潘峰等[15]的方法测定菌株的抗氧化活性(总抗氧化活性测定(ABTS 法)和DPPH 自由基清活性试验);采用平板对峙法测定菌株对大肠杆菌、金黄色葡萄球菌、枯草芽孢杆菌、米根霉和橘青霉的抑制功能[16];参考胡宇玲等[17]方法测定菌株产淀粉酶、蛋白酶、脂肪酶、果胶酶的功能.
2 结果与分析
2.1 不同季节油樟茎内生真菌的分离及群落组成
通过培养技术共获得98 株内生真菌,其中春、夏、秋、冬季分别分离得到27、20、28、23 株菌,秋季获得的内生真菌数量最多,占内生真菌总菌株数的28.57%.由表1 可知,春季油樟茎分离的27 株内生真菌隶属于13 个属22 种,其中附球菌属(Epicoccum)与链格孢属(Alternaria)为春季油樟茎中的优势菌属.春季分离菌株的系统发育树见图1.由表2 可知,夏季油樟茎获得的20 株内生真菌隶属于8 属19种,其中Diaporthe为第1 优势菌属,Alternaria与Neofusicoccum均为第2 优势菌属,夏季分离菌株的系统发育树如图2所示.由表3可知,秋季油樟茎分离的28株内生真菌隶属于13个属22种,其中Alternaria为优势菌属.秋季分离菌株的系统发育树见图3.由表4可知冬季油樟茎分离的23株内生真菌隶属于14 个属22 种,其中Alternaria为优势菌属,冬季分离菌株的系统发育树如图4所示.

图1 基于ITS序列构建的春季油樟茎内生真菌系统发育树

图2 基于ITS序列构建的夏季油樟茎内生真菌系统发育树

图3 基于ITS序列构建的秋季油樟茎内生真菌系统发育树

图4 基于ITS序列构建的冬季油樟茎内生真菌系统发育树
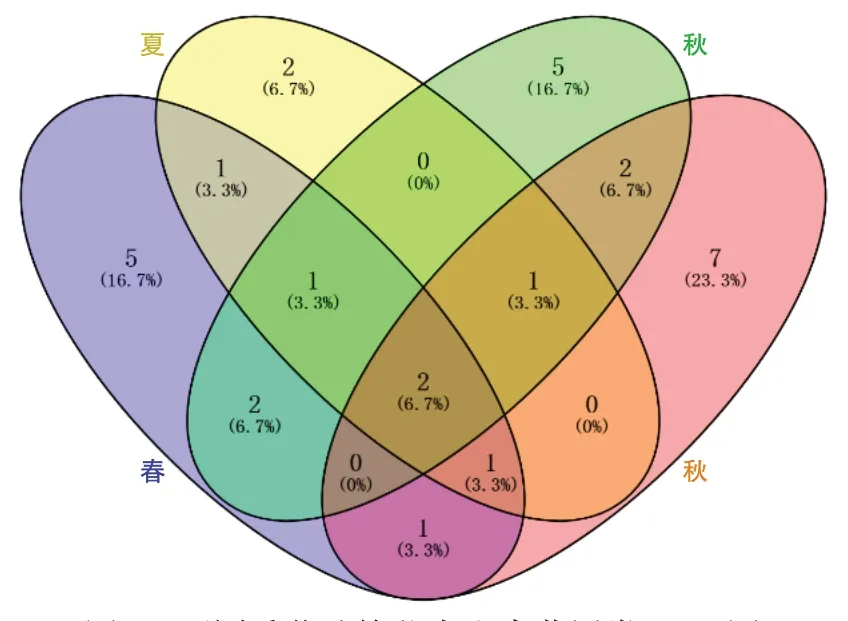
图5 不同季节油樟茎内生真菌属类Venn图
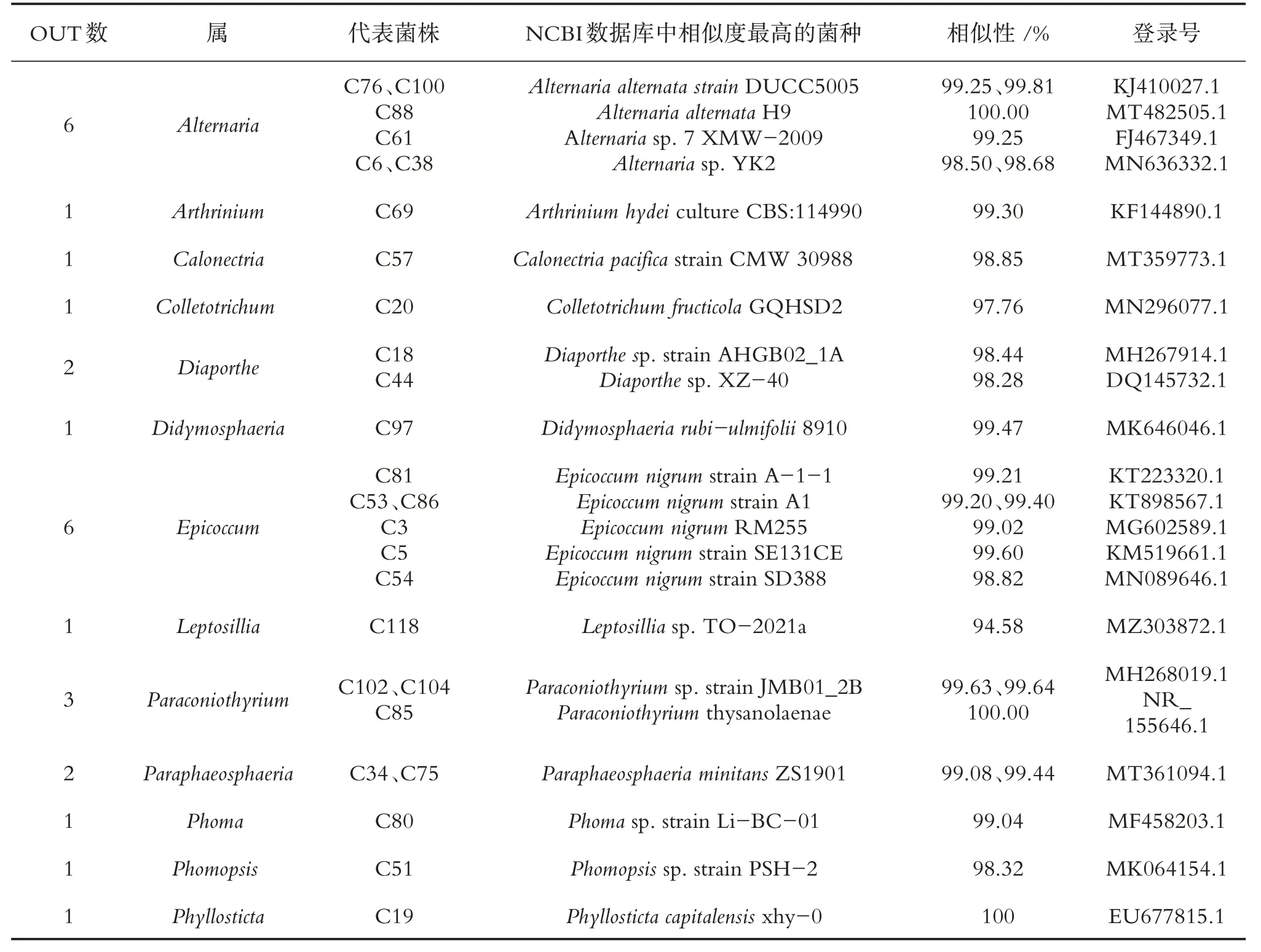
表1 春季油樟茎内生真菌分布
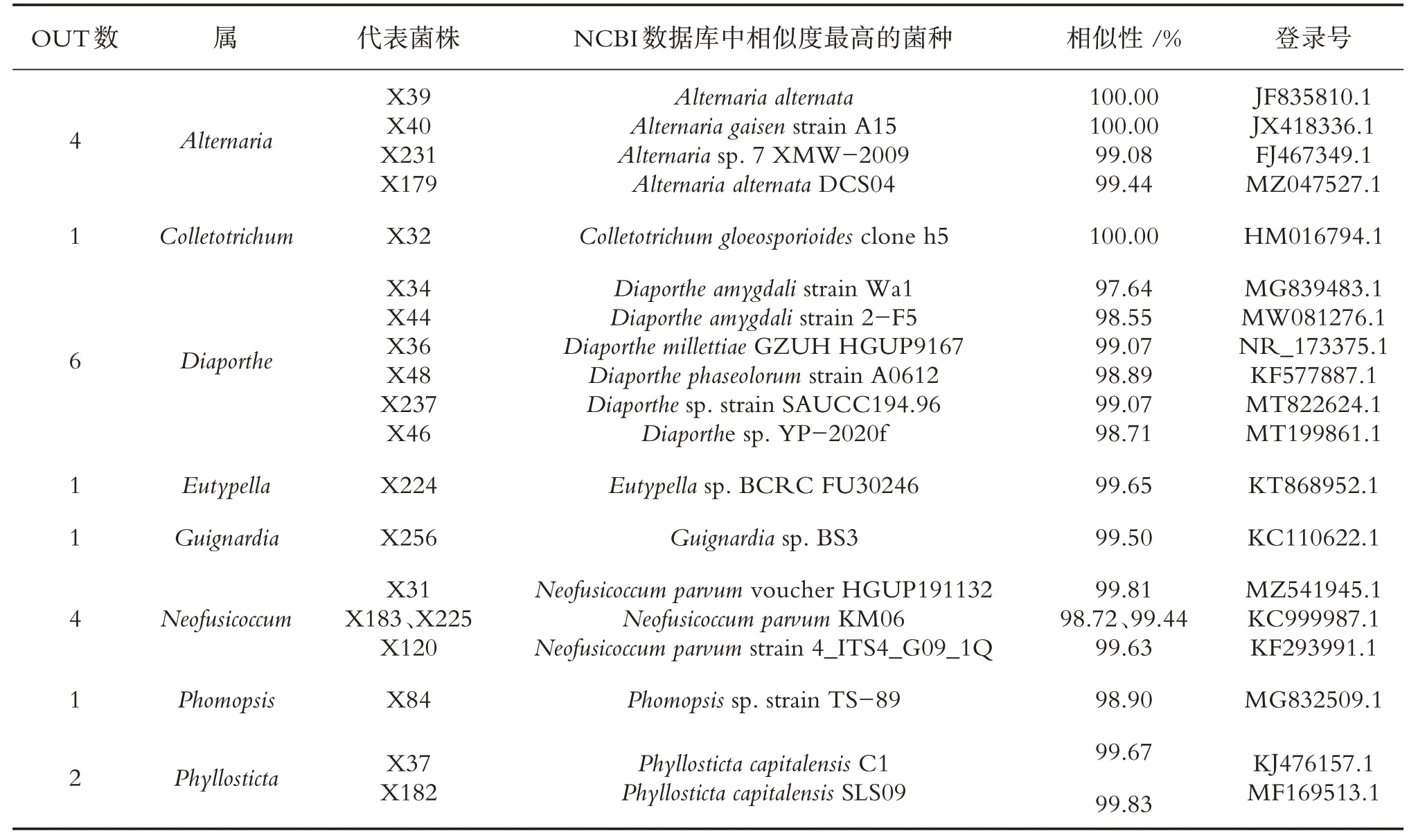
表2 夏季油樟茎内生真菌分布
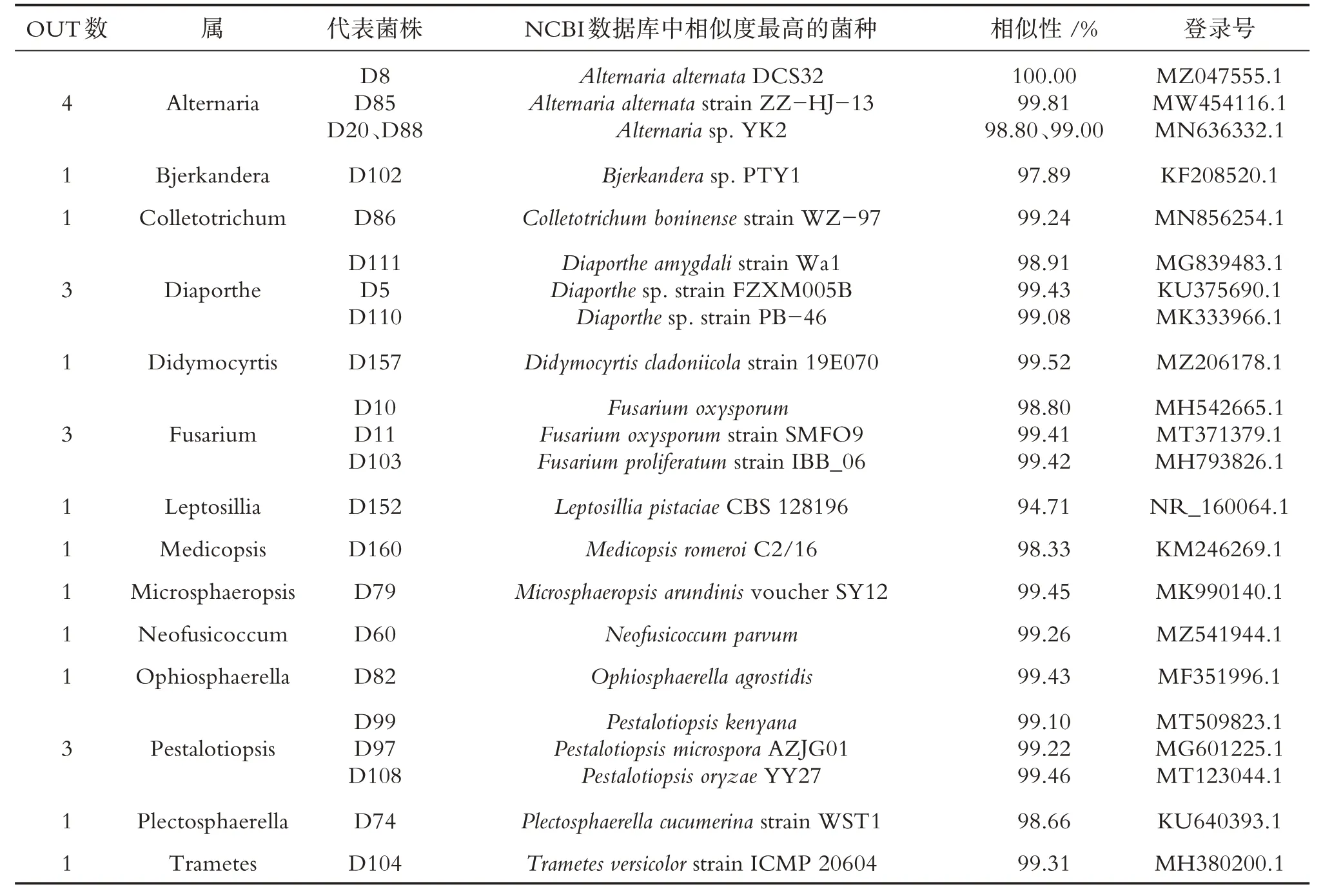
表4 冬季油樟茎内生真菌分布
对油樟茎部内生真菌进行生物分类学统计可知,98 株内生真菌隶属于2 门3 纲8 目21 科30 属,子囊菌门(Ascomycota)为优势门,座囊菌纲(Dothideomycetes)为优势纲,格孢腔菌目(Pleosporales)为优势目,格孢菌科(Pleosporineae)为优势科,Alternaria为优势属.春季27 株内生真菌隶属于1 门2 纲6 目10科13属,夏季20株内生真菌隶属于1门2纲6目7科8属19种,秋季28株内生真菌隶属于1门2纲5目11科13 个属22 种,冬季23 株内生真菌隶属于2 门3 纲7 目12 科14 个属22 种.由图1 可知,4 个季节共有属2 个,为Alternaria和Colletotrichum.春、夏、秋、冬特有属分别为5、2、5、7 个,其中Arthrinium、Calonectria、Epicoccum、Paraphaeosphaeria、Phoma为春季特有属;Eutypella、Guignardia为夏季特有属;Austropleospora、Didymella、Periconia、Shiraia、Pyrenochaetopsis为秋季特有属;Bjerkandera、Didymocyrtis、Microsphaeropsis、Ophiosphaerella、Pestalotiopsis、Plectosphaerella和Trametes为冬季特有属.
2.2 不同季节油樟茎内生真菌的多样性
香农-维纳指数H'和辛普森指数D均反映物种的丰富度,均匀度指数E用于分析内生真菌群落分布的均匀程度,通过表6 的多样性分析可知,不同季节油樟茎内生真菌S指数、H'指数、D指数和均匀度指数E均表现为: 冬季>春季 ≈ 秋季>夏季,因此,冬季油樟茎内生真菌群落的多样性相对最丰富,夏季最低.
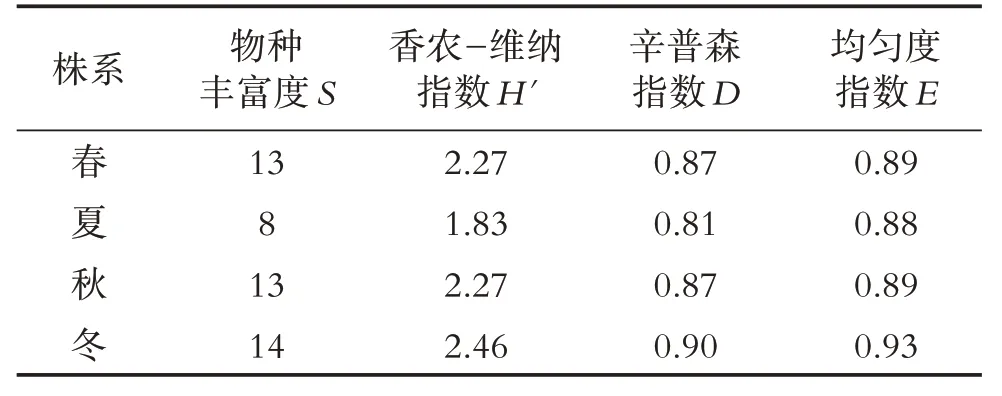
表6 不同季节油樟茎内生真菌的多样性指数
2.3 内生真菌产黄酮功能筛选
通过显色反应初筛,98 株内生真菌中Q73、C18、Q32、Q13、D20、C5、C80、X32、X44、X120、X224、X225、C34、D79 和D97 菌株具有较好的产黄酮能力, 其中4 株分离于春季, 5 株分离于夏季,3 株分离于秋季, 3 株分离于冬季, 共归类于1 门2纲5 目8 科11 属,分别是Alternaria、Microsphaeropsis、Epicoccum、Phoma、Paraconiothyrium、Colletotrichum、Diaporthe、Eutypella、Pestalotiopsis、Paraphaeosphaeria和Neofusicoccum. 其中Q73 显色反应最好,属于Alternaria,4 种显色试剂均显色.通过紫外分光光度计法测定可得Q73 菌丝体产黄酮量3.29 mg/g,发酵液产黄酮量为0.04 mg/mL.
2.4 Q73内生真菌其他功能的筛选
ABTS 自由基清除试验通常用于评价化合物总抗氧化活性,DPPH 是一种稳定自由基组成的深蓝色结晶粉末,通常用于天然产物对自由基清除活性的评价.由实验结果可以看出,菌株Q73 均具有较好的抗氧化活性,其中菌丝体ABTS 的清除活性为(72.93±2.31)%,发酵液ABTS 清除率为(56.00±0.98)%,对DPPH 清除活性分别为(82.60±1.58)%、(27.68±0.63)%.在多酚含量测定中可得,Q73 菌株菌丝体多酚含量为2.114 mg/g,发酵液多酚含量为104.82 ug/ml.通过产酶功能筛选发现,Q73 在果胶酶鉴别培养基上产生了光晕,说明Q73 具有产果胶酶的功能,其他鉴别培养基上未发现透明圈.在抑菌功能测定中显示,Q73菌株对枯草芽孢杆菌、大肠杆菌具有较强的抑菌活性,对其他病原菌无拮抗作用.
3 讨论
内生真菌是参与宿主植物生长、发育及代谢等过程的重要因子,极具研究价值并涵盖丰富的微生物资源,为开发潜在菌株并分析与宿主功能相关性具有重要意义,其定殖于植物体内受到内部因子(宿主植物种类)和外部因子(气候条件、地理位置、温度等)因素的影响[18].目前对植物茎中内生细菌多样性的研究较多,而内生真菌相对较少.辛赫文等[19]从河南小麦茎部分离获得100 株内生真菌,其中子囊菌是小麦内生真菌的主要类群,优势属为Penicillium,相对分离频率为66.70%.宁祎等[20]对桃儿七茎部内生真菌研究也发现,子囊菌门占绝对优势,但优势属为拟青霉属.本研究从不同季节油樟茎组织共获得98株内生真菌,隶属于2门3纲8目21科30 属,其中优势门为Ascomycota,优势属为Alternaria.优势门与上述研究相同,但优势属存在差异,这正是因为宿主植物种类不同而呈现不同的内生真菌组成.Alternaria隶属于真菌界、子囊菌门、座囊菌纲、格孢腔菌目、格孢菌科,广泛存在于禾本植物中[21],研究表明该属类大多数菌种寄生在植物中,少数菌株可侵染人和动物,同时发现Alternaria具有应用潜力的生物资源,如长柄链格孢部分菌株的分生孢子可制备生防制剂,用于防治烟草的赤星病[22],且链孢霉菌丝体能提取得到蛋白质、糖分、淀粉等产物,用于饲料中可提高营养价值,促进精饲料的形成[23].Shahid 等[24]研究表明Alternaria、Epicoccum和Fusarium在许多植物组织的定殖率均最高,与本试验相同,从油樟茎中均分离到以上属类菌株,但相比于宏基因组技术,利用传统组织分离法无法分离到油樟茎中不可培养的内生真菌,因此后续还将利用高通量技术实现茎部内生真菌的分析.
按季节看,春季27 株内生真菌隶属于1 门2 纲6目10 科13 属,夏季20 株内生真菌隶属于1 门2 纲6目7科8属19种,秋季28株内生真菌隶属于1门2纲5 目11 科13 个属22 种,冬季23 株内生真菌隶属于2门3 纲7 目12 科14 个属22 种.通过多样性指数和均匀度结果发现内生真菌数量和种属随季节不同而不同,冬季油樟茎内生真菌群落的多样性相对最丰富,夏季最低.吕立新[25]对不同季节茅苍术内生真菌群落多样性的研究发现,内生真菌群落会随着季节变化呈现出一定演化规律,其中夏季内生真菌的多样性高于春季和秋季.罗鑫等[26]采用组织分离法对30 年生健康马尾松不同季节内生真菌的分离,表明秋季马尾松内生真菌最为丰富,其次为夏季,而冬季与夏季相似.崔文霞等[27]对不同季节鱼腥草内生真菌分离中发现,交链孢属Alternaria和镰孢菌属Fusarium为优势菌群,且夏季分离得到的内生真菌最多.与本试验结果不同,说明不同季节内生真菌的差异性可能与菌株对营养需求的特殊性有关[28],且不同植物内生真菌存在季节性变化的差异.
植物内生真菌对植物的生长具有积极作用,并能代谢产生与宿主相同或相似的天然产物[3].研究发现在野生大豆内生真菌Y2-2 和柽柳内生真菌菌株C1T2-4 菌株的代谢产物中可检测出脂肪酸[29];Yang 等[30]从大豆中分离出了一株脂质含量高达56%、脂肪酸甲酯含量高达98%的内生菌株,且可用作生产生物柴油;Jagannath 等[31]发现从斑籽根、叶、茎、花和种子组织中分离的内生真菌具有产纤维素酶的功能.关于油樟的研究主要集中在活性成分提取、组织栽培和成分的功能性研究,对内生菌的研究也多数局限于内生细菌,因此,油樟茎部内生真菌生物活性代谢产物的研究具有较大开发利用空间.目前有麻花艽[32]、青钱柳[33]、番木瓜[11]分离内生真菌筛选产黄酮功能,且产黄酮能力各有差异.青钱柳[33]中内生真菌Alternaria产黄酮含量为3.61 mg/L;明日叶MRY-1 内生真菌菌丝体黄酮产量为1.194 mg/g[34];荷叶中筛选的棘孢曲霉Aspergillusaculeatus内生菌总黄酮为8.2 mg/g[35],本文筛选得到Q73 菌丝产黄酮量3.29 mg/g,发酵液产黄酮量为0.04 mg/mL,说明Q73 产黄酮能力中等,后期将进一步优化黄酮类物质的提取方法,提高黄酮获得量,同时发现发酵液活性物质的产量低于菌丝体含量,对菌丝体和发酵液中多酚物质的测定也证明了此结论.
据报道一定自由基对于维持正常生命活动所必需,但若自由基过多会加速机体衰老,诱发人类心脑血管疾病、肿瘤、阿尔茨海默等多种疾病的发生[36],因此天然高效的抗氧化剂越发受到重视.本试验通过菌株菌丝体和发酵液的ABTS 和DPPH 抗氧化能力的测定,发现其抗氧化能力与黄酮和多酚含量成正比,与其他研究结果相同,潘峰[15]证实不同溶剂提取物的黄酮和多酚含量与抗氧化活性成正相关关系;Ye 等[37]从银杏(Ginkgo biloba)分离获得一株内生真菌ChaetomiumglobosumCDW7 能产黄酮且具有很强抗氧化活性,同时显示黄酮物质为菌株主要的抗氧化活性成分.本试验还测定了Q73菌株具有产果胶酶、抗枯草芽孢杆菌和抗大肠杆菌的能力,值得后续进一步研究.
目前国内外还未见关于油樟茎内生真菌多样性的研究,大量的内生真菌资源还有待挖掘和开发,为了更好地研究油樟茎内生真菌种群多样性,有必要采集更多不同地区样品,以获得更多可培养的内生真菌,从而为今后深入探讨不同地域油樟茎内生真菌多样性提供理论依据,也为进一步阐明油樟茎与内生真菌相互关系、发掘活性菌株奠定基础,同时为生物活性物质筛选和油樟资源的开发提供依据.
猜你喜欢
绿色天府(2022年2期)2022-03-16
广州化工(2021年22期)2021-11-30
湖北农机化(2020年4期)2020-07-24
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
乡村地理(2018年2期)2018-09-19
兵器知识(2018年5期)2018-05-30
四川林业科技(2015年6期)2015-12-02
宜宾学院学报(2014年12期)2014-07-20
集美大学学报(教育科学版)(2014年2期)2014-02-28
