
NO对铝胁迫下绛三叶种子萌发及幼苗生长的影响
2023-12-21 15:35周万海胡连清刘雯雯蔡意凤冯瑞章魏琴赵鑫陈露
宜宾学院学报 2023年12期
周万海,胡连清,刘雯雯,蔡意凤,冯瑞章,魏琴,赵鑫,陈露
(1.宜宾学院 农林与食品工程学部,四川宜宾 644000;2.香料植物资源开发与利用四川省高校重点实验室,四川宜宾 644000;3.自贡市农业科学研究院,四川自贡 643000)
豆科植物(Legμminosae)因富含油脂、蛋白质、 淀粉和维生素等营养物质,具有重要的社会和经济意义[1].近年来,由于酸雨和酸性肥料的使用,土壤酸化趋势逐步增加,严重影响豆科作物的生长和产量.酸化土壤中,限制作物生长和产量的因素多种多样[2],除氮(N)、磷(P)等必需营养元素外,铝(Al)影响最大,是限制作物生产的最关键因素之一,研究表明,酸化土壤中铝常以不同形式存在,其中离子态[Al3+、Al(OH)2+]对植物毒害最重,铝毒对植物的伤害首先表现于破坏根尖结构,抑制根系生长,进而影响植物的产量与品质[3].一氧化氮(NO)是一种普遍存在于生物体中的信号分子,广泛参与生物体内各种反应,在植物生长发育和抗逆反应中起关键作用[4].一定浓度的NO 对植物侧根、不定根和根毛的发育有促进作用[5];盐胁迫下外源添加亚硝基铁氰化钠(SNP)提高了苜蓿根系活力,促进了苜蓿幼苗根系生长[6];外源添加SNP 能减轻铝胁迫下西瓜根系氧化损伤,并通过影响根系对营养元素的吸收来缓解铝毒作用[7-8].因此,研究NO 对植物抗逆性的调控效应及作用机制不仅具有重要的理论意义和实践价值,而且对保护农业生态系统和探究受损环境的生物修复途径也具有重要意义.
豆科饲草绛三叶(T.incarnatum)为车轴草属一年生优质牧草,因茎叶富含蛋白质且还可与根瘤菌共生固氮来促进自身生长和提高土壤肥力,目前已成为南方地区冬闲田的主栽牧草之一[9-10],但受土壤酸化影响,铝毒害已成为限制绛三叶生产的主要土壤环境因子之一,目前有关NO 对绛三叶耐铝的缓解效应研究未有报道.为进一步阐明NO 在铝中的作用,本文拟从豆科植物适应铝胁迫的限度以及探究外源NO 对铝胁迫下的豆科作物生理影响出发,探究绛三叶耐铝毒害的能力以及外源NO 缓解铝胁迫的机制,为进一步理解铝胁迫对豆科植物生长抑制的作用机制及调控方法提供依据.
1 材料与方法
1.1 试验材料
绛三叶种子由河南世纪天缘生态科技有限公司提供,外源NO 供体为亚硝基铁氰化钠(SNP),铝离子来源为分析纯AlCl3.
1.2 试验方法
①铝胁迫对绛三叶种子萌发的影响.配置试验处理液: CK(无菌水)、500、1 000、2 000、3 000、3 500、4 000、5 000、10 000 μM 的AlCl3溶液(均含有0.5 mM 的CaCl2溶液,pH4.0),选取饱满均匀的绛三叶种子均匀置于垫有2 层滤纸的培养皿中,加入20 mL 处理液,每个处理组3 次重复,然后将培养皿置于25℃,12 h 光照/12 h 黑暗,湿度为60%的恒温培养箱中发芽生长,处理4 d 后测定发芽势,处理10 d后测定根长、芽长、生物量和发芽率.将该试验结果得出的适宜铝胁迫处理浓度记录为FA.
②外源NO 对铝胁迫下绛三叶种子萌发的影响.根据方法①的结果选取适宜的铝胁迫处理浓度FA.试验设置CK(无菌水)、FA、FA+100 μmol/L SNP、FA+150 μmol/L SNP、FA+200 μmol/L SNP、FA+300 μmol/L SNP 共6 个处理,选取均匀饱满无损伤的种子,每皿放置50 粒,加入20 ml 处理液,每个处理3 次重复,之后每两天加5 ml 处理液,10 d 后测定发芽率、侧根数、主根长、芽长以及生物量.根据该结果选取添加外源SNP的适宜浓度记录为FB.
③外源NO 对绛三叶幼苗生长及理化特性的影响.根据方法①②的结果采用沙培法,选取饱满均匀的种子,消毒后播种于营养钵,发芽后,每3 d浇灌一次GS营养液,生长40 d后进行铝胁迫处理.试验设置如下处理:CK(Hoagland 营养液)、FB、FA、FA+FB 溶液4 个处理,每组处理重复3 次,培养条件为24 ℃/14 h,夜间20 ℃/10 h,相对湿度为60%~70%,处理10天后,剪取真叶,分装于锡箔纸袋中,用液氮速冻后置于超低温冰箱保存,用于后续理化指标测定.
1.3 测定指标与方法
(1)发芽率和发芽势的测定.发芽势:绛三叶种子萌发第4 d测定,以露出种皮的胚根长度达到种子自身长度为发芽的标准,以百分比表示,即发芽势(%) = 4 d发芽的种子数/供试种子数×100%.
发芽率:绛三叶种子萌发10 d后测定,以突出种皮的胚根长度达到种子自身长度为发芽的标准,以百分比表示,即发芽率(%) = 10 d 发芽种子数/供试种子总数×100%.
(2)主根、芽、侧根及生物量指标测定.每个试验处理组随机抽取10 株幼苗,用游标卡尺分别测量其主根长、胚芽长、侧根数量,最后取平均值.从每个试验处理组中随机抽取20 株幼苗,在分析天平上称重,取其平均值得到每株幼苗的鲜重;再在105 ℃下杀青15 min,恒温烘箱中75 ℃条件下烘干1 h;最后称重,取平均值得到每株幼苗干重.
(3)生理指标测定.叶绿素定量测定采用乙醇提取比色法;可溶性蛋白含量测定采用考马斯亮蓝G-250 方法[11];膜脂过氧化产物丙二醛(MDA)含量测定采用硫代巴比妥酸法[11];羟自由基含量测定采用Halliwell 的方法[12];过氧化氢酶(Catalase,CAT)活性测定参考Beers和Sizer的方法[13];抗坏血酸过氧化物酶(APX)活性测定参考Nakano and Asada 的方法[14];愈创木酚过氧化物酶(GPX)活性测定参考Mrbanek的方法[15].
1.4 试验数据处理
用Excel 2010 软件与Origin 2018 软件处理数据和绘图.
2 结果与分析
2.1 铝胁迫对绛三叶种子萌发生长状况影响
由表1 可知,随铝离子浓度的增加,绛三叶种子萌发逐步受到抑制,Al3+浓度升高到2000 μmol/L时,绛三叶种子的发芽势与对照无差异,但发芽率比对照降低4.0%,根芽长、鲜重和干重分别降低2.942 cm、0.694 g和0.013 g,表明绛三叶受到铝毒害,因此后期试验选取其为铝胁迫处理浓度FA.当Al3+浓度为3500~5000 μmol/L 时,绛三叶发芽速度明显缓慢,发芽率显著下降;5000 μmol/L 处理下,发芽率比对照降低40%.
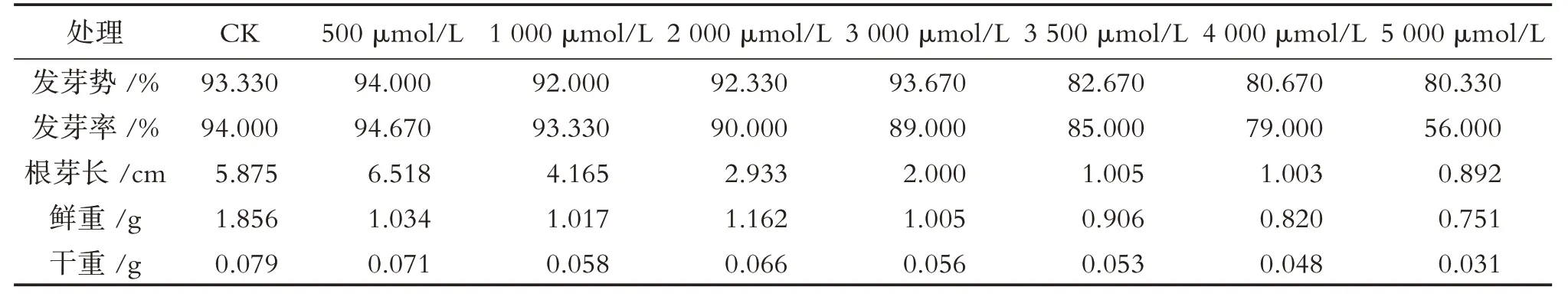
表1 不同浓度酸铝胁迫下绛三叶生长状况的影响Tab.1 Effects of different concentrations of aluminum stress on the growth of T. incarnatum
2.2 外源NO对酸铝胁迫下绛三叶种子萌发的影响
由表2可知,在2 000 μmol/L Al3+胁迫下绛三叶种子萌发的各项生长指标明显受到抑制,其幼苗干重、鲜重、根系以及胚芽长与CK 相比明显降低.而在2 000 μmol/L Al3+胁迫下添加外源NO后,绛三叶受铝胁迫作用明显缓解,外源添加100 μmol/L 与150 μmol/L SNP,绛三叶种子的发芽率与侧根数量升高;从绛三叶幼苗根系及芽长来看,铝胁迫明显抑制绛三叶种子萌发后的根系与芽的生长,与对照(单一2 000 μmol/L Al3+胁迫)相比,添加100 μmol/L、150 μmol/L 和200 μmol/L 的SNP 提高了绛三叶种子萌发后的根芽的生长指标.从绛三叶幼苗干、鲜重分析,添加100 μmol/L SNP 提高了绛三叶幼苗的干重与鲜重.综合来看,外源添加100 μmol/L SNP处理对铝胁迫下绛三叶的促生效应最明显,后期试验采用此添加外源NO处理方式(FB).
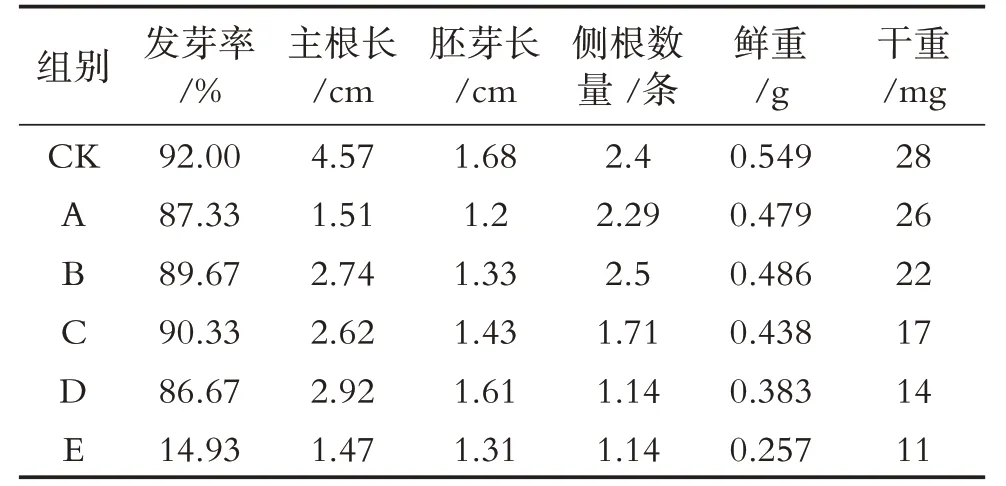
表2 SNP对铝胁迫下绛三叶生长状况的影响Tab.2 Effect of SNP on growth condition of T. incarnatum under aluminum stress
2.3 外源NO对铝胁迫下绛三叶幼苗理化特性的影响
(1)外源NO 对绛三叶幼苗叶绿素及类胡萝卜素含量影响.由图1 可知,单独用100 μmol/L SNP处理的实验组,叶片的叶绿素a、b 和类胡萝卜素含量无明显变化.2 000 μmol/L-1Al3+处理下与CK 相比绛三叶幼苗叶片中叶绿素a、叶绿素b 与类胡萝卜素的含量分别下降了15.8%、19.6%、18.8%.2 000 μmol/L Al3+处理下添加100 μmol/L SNP,与单一铝胁迫处理相比,绛三叶幼苗叶片中叶绿素a 含量上升了13%;叶绿素b 的含量上升了6.5%;类胡萝卜素含量上升了7.7%.结果表明,在铝胁迫下,绛三叶幼苗叶片中的叶绿素与类胡萝卜素含量降低.在添加SNP 后绛三叶幼苗叶片中的叶绿素和类胡萝卜素含量有明显升高,表明NO 对铝胁迫造成的光合色素降解有一定的缓解效应.

图1 SNP对绛三叶幼苗色素含量的影响Fig.1 Effect of SNP on pigment content of T. incarnatum seedling under aluminum stress
(2)外源NO 对绛三叶幼苗叶片可溶性蛋白含量的影响.单独用100 μmol/L SNP 处理的实验组,叶片的可溶性蛋白含量与空白对照组CK 差异不大,2 000 μmol/L Al3+溶液处理使绛三叶幼苗叶片中可溶性蛋白含量上升了27.6%,在此条件下添加100 μmol/L 的SNP 溶液后,与单一用Al3+处理的绛三叶相比,叶片中可溶性蛋白含量下降了15.8% (图2).
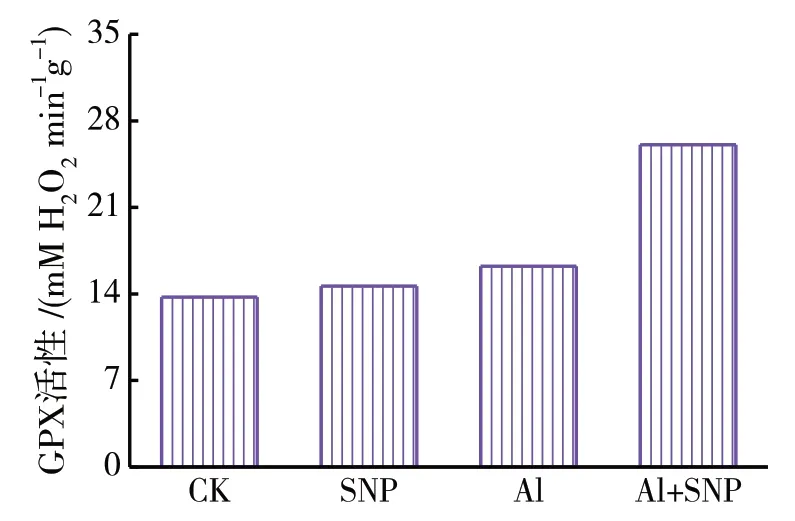
图2 SNP对绛三叶幼苗可溶性蛋白含量的影响Fig.2 Effect of SNP on soluble protein content of T. incarnatum seedling under aluminum stress
(3)外源NO 对绛三叶幼苗叶片羟自由基含量的影响.用100 μmol/L SNP 处理的叶片中羟自由基含量与CK 相比差别不明显,在铝胁迫下绛三叶幼苗叶片中羟自由基与CK 相比增加了103.5%,在添加100 μM SNP 后绛三叶幼苗叶片内羟自由基含量下降了31%(图3).这表明铝胁迫会使绛三叶幼苗叶片中羟自由基大量累积,外源添加NO 能缓解铝胁迫对绛三叶造成的毒害.
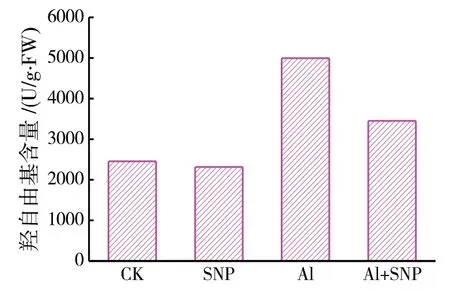
图3 SNP对绛三叶叶片羟自由基含量的影响Fig.3 Effect of SNP on Hydroxyl radical content of Crimson T. incarnatum under aluminum stress
(4)外源NO 对绛三叶幼苗叶片抗氧化酶活性的影响.由图4 可知,单独用100μmol/L SNP 处理的实验组与空白对照组CK 相比,叶片中CAT 活性降低不明显,GPX 和APX 活性分别上升了6.9%和13.7%.与CK 相比在铝胁迫下绛三叶幼苗中GPX活性上升了18.3%,CAT 和APX 活性分别下降了24.7%、51.4%;在添加SNP 溶液处理后,绛三叶幼苗叶片中,CAT、GPX、APX 分别升高了60.4%、60.5%、166.2%.结果表明,铝胁迫能引起绛三叶氧化反应和酶系统活性改变,而外源NO 对铝胁迫下绛三叶幼苗的抗氧化系统有一定的激活作用,能增加绛三叶抗氧化酶活性,缓解氧化胁迫对植物造成的伤害.

图4 SNP对绛三叶幼苗叶片抗氧化酶活性的影响Fig.4 Effects of SNP on activities of APX、GPX and CAT in T. incarnatum leaf under aluminum stress
(5)外源NO 对绛三叶幼苗叶片膜脂过氧化产物(MDA)含量的影响.由图5 可知,单独用100 μmol/L SNP 处理的实验组,叶片MDA 含量比CK降低16.2%.铝胁迫下绛三叶植株叶片MDA 含量则比CK 增加17.8%.与单一铝胁迫处理相比,添加100 μmol/L SNP,降三叶幼苗中膜脂过氧化产物含量下降了12.1%.这表明铝胁迫会致使绛三叶片膜脂过氧化程度加深,而外源添加SNP 能缓解铝对植物造成的毒害.

图5 SNP对绛三叶叶片MDA含量的影响Fig.5 Effect of SNP on MDA content of T. incarnatum leaf under aluminum stress
3 讨论
南方地区酸性土壤中富含大量的活性铝离子[16],抑制作物生长发育,影响作物品质及产量,对作物产生严重毒害作用.在实际生产中,铝毒对作物生产最初的影响是种子萌发,梅文君等[17]发现在酸胁迫下,楚雄南苜蓿种子的发芽率、发芽势等都随着pH 值的降低而降低,在强酸性环境下,楚雄南苜蓿幼苗根长、茎长等显著低于其他处理;许光德等[18]发现铝胁迫明显对向日葵产生毒害作用,促使细胞膜脂过氧化加重,抑制了种子的萌发以及幼苗的生长;张雅茹等[19]发现铝胁迫使油菜幼苗的生长受到明显的抑制,油菜的生物量降低.研究表明,添加适量的外源NO 可以有效提高植物种子的发芽率、发芽势、幼苗根长和茎长[20];适当浓度的外源NO 可以有效促进萝卜种子萌发,减弱铝胁迫下对萝卜幼苗根长、茎长、叶长、叶宽产生的副作用[21];张汉林等[22]发现外源NO 能够促进铝胁迫下黑麦草的生长和生物量的积累.本研究中,较低浓度的铝胁迫,对绛三叶种子萌发及生长影响不大,表明其有一定程度的耐铝能力,但高浓度铝胁迫处理则明显抑制了绛三叶种子萌发,且明显降低了种子的发芽率和幼苗生物量积累,这与前人研究结果相一致[8,23-24];在铝胁迫下对降三叶外源添加100 mM SNP,其发芽率以及幼苗鲜、干重增加,说明NO 可以缓解铝毒对绛三叶种子萌发和幼苗生长的抑制.
叶绿素参与植物光能的吸收、传递和转化,类胡萝卜素参与植物光能的吸收、传递,二者是反映植物光合能力的一个重要指标.光合作用为植物进行碳同化、有机物积累、为植物生长发育提供能量的过程[25].铝离子会破坏植物的光合系统,降低叶绿素和类胡萝卜素的含量,抑制植物体的光合作用[26].研究表明,铝胁迫下西瓜叶片的叶绿素含量明显降低,西瓜的生长与产量受到较大的影响[8].适量的外源NO 可增加植物体内叶绿素的含量,从而增强植物的光合作用[27].有研究表明,外源NO 处理可显著增加黑麦草幼苗叶绿素a 的含量,提高叶绿素总含量[22].本试验中,高铝胁迫下(2 000 μmol/L)绛三叶叶绿素与类胡萝卜素含量明显降低,光合色素的降低会直接影响植物的光合能力,从而影响植物的生长,降低作物的产量;添加一定外源NO 促进了绛三叶叶绿素与类胡萝卜素的合成,从而提高其光合作用能力,促进其生长,增加有机物的积累.该结果说明外源NO 可以逆转高铝胁迫下绛三叶光合作用能力的降低,并缓解铝对作物的毒害作用.
逆境胁迫下植物体内会产生大量的活性氧自由基并积累,破坏活性氧自由基与抗氧化酶之间的代谢平衡,若活性氧不能及时被清除,则会引起细胞膜系统受损,从而影响植物生理状态和生长发育,植物膜脂过氧化过程中还会产生一些渗透调节物质,胁迫程度越高,其含量越高[23].CAT、GPX、APX 等抗氧化酶能够有效地清除活性氧[28].NO 为调节植物生长发育的重要信号分子,其具有抗氧化作用,并能阻断脂质过氧化反应、保护植株免受超氧化损伤以及增强植物细胞中抗氧化酶活性,从而缓解金属离子对植物造成的毒害作用[29].研究表明,NO 可通过提高渗透调节物质以及抗氧化能力来增强植物的抗逆性,同时显著降低MDA 含量,缓解逆境造成的损伤[30].在铝胁迫下萝卜幼苗叶片中的CAT 和APX 活性显著下降,根尖的O-2产生速率和H2O2含量显著上[21].肖家昶[8]等发现铝胁迫下西瓜根系可溶性糖与脯氨酸含量显著上升,可溶性蛋白的含量无显著差异,CAT 等抗氧化酶活性显著降低,MDA含量增加,添加外源NO 增加了铝胁迫下西瓜根系中脯氨酸含量显著积累,促进西瓜根系抗氧化酶的合成,从而减少活性氧损伤,缓解铝胁迫下生理损伤;铝胁迫下外源添加NO 西瓜幼苗的CAT 等抗氧化酶活性及渗透调节物质含量提高,MDA 减少,从而减轻质膜过氧化的损伤[23].本试验中,铝胁迫下绛三叶幼苗和可溶性蛋白含量升高,CAT、GPX、APX 活性明显降低,膜脂过氧化产物MDA 明显积累,该结果表明绛三叶受到严重铝毒害;在添加100 mM SNP 后,绛三叶幼苗叶片中CAT、GPX 和APX抗氧化酶活性明显提高,羟自由基含量和膜脂过氧化产物MDA 积累明显降低,从而缓解铝胁迫下绛三叶幼苗的生理损伤,该结果与前人研究一致[8,23,30].但外源添加NO 后可溶性蛋白的含量降低,这可能是因降三叶幼苗是通过调节自身其他渗透调节物质抵抗逆境胁迫所致.
本研究表明,在2 000 μML 铝胁迫下对绛三叶种子进行添加100 μM NO 处理,其光合色素含量提高,抗氧化酶活性提高,膜脂过氧化程度降低,其生长指标变好,绛三叶能更好地生长.结果表明,添加NO 提高了绛三叶对铝胁迫的适应能力,缓解了铝胁迫对绛三叶的毒害作用.该结果能为进一步理解铝胁迫对豆科植物生长抑制的作用机制及调控方法提供参考依据.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
阅读(科学探秘)(2020年8期)2020-11-06
西藏农业科技(2019年3期)2019-11-04
中国果业信息(2019年1期)2019-01-05
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
现代园艺(2018年3期)2018-02-10
生物学教学(2017年9期)2017-08-20
上海农业学报(2017年3期)2017-04-10
天津农林科技(2015年1期)2015-12-30
