
山奈酚通过调控AMPK/Nrf2/HO-1信号通路缓解ox-LDL介导的内皮细胞损伤
2018-04-20 02:29康桂兰景增秀
中国免疫学杂志 2018年4期
康桂兰 景增秀
(西宁市第二人民医院,西宁 810003)
血管内皮细胞是维持血管壁正常结构和功能的重要屏障,内皮损伤是动脉粥样硬化(Atheroscle-rosis,AS)的始动环节[1]。有文献报道,氧化型低密度脂蛋白(oxidized low-density lipopro-tein,ox-LDL)是早期AS发生和发展的重要因素,在AS病灶部位中存在明显增加的氧化的低密度脂蛋白[2,3]。因此减少ox-LDL对内皮细胞的氧化损伤是预防和治疗AS的有效途径。黄酮类化合物是天然抗氧化剂,能够缓解脂蛋白的过氧化作用,进而有效地防治AS的发生[4]。山萘酚(Kaempferol)是一种黄酮醇类化合物,广泛存在于水果、蔬菜、豆类、茶叶中,具有抗癌、抗炎、抗氧化等多种生理作用[5,6]。已有研究表明,山奈酚能够抵抗缺血再灌注等心血管疾病损伤[7,8],抵抗AS[9],但是具体的分子机制并不明确。本研究在体外通过用ox-LDL诱导人脐静脉内皮细胞(Human umbilical vein endothelial cells,HUVECs)损伤,进一步探讨山奈酚对ox-LDL诱导的细胞增殖、凋亡、炎性因子和黏附分子产生和氧化应激的影响,并探讨其作用机制。
1 材料与方法
1.1材料 HUVECs购自ATCC;山奈酚购自Sigma公司;MTT试剂盒、annexin V-FITC/PI细胞凋亡试剂盒购自南京凯基生物有限公司;超氧化物歧化酶(Superoxide dismutase,SOD)试剂盒购于南京建成生物工程研究所;核提取试剂盒购于Pierce 公司;RIPA 细胞裂解液、PMSF和BCA 蛋白含量测定试剂购自上海碧云天生物技术研究所;Anti-cleaved-caspase-3、anti-Bcl-2、anti-TNF-α、anti-IL-1β、anti-IL-6、anti-VCAM1、anti-ICAM1、anti-E-selectin、anti-p-AMPK、anti-Nrf2、anti-HO-1和anti-GAPDH等 I 抗购自Abcam公司和Santa Cruz 公司;Ⅱ抗购自北京中杉金桥生物有限公司。
1.2方法
1.2.1细胞培养及药物处理 将HUVECs置于含有10%胎牛血清的DMEM培养基中,在37℃、5% CO2的培养箱中培养。当细胞生长至80%~90%融合时,用0.25%的胰酶进行消化传代,分别接种到不同规格的培养板中。药物处理:先用山奈酚预处理HUVECs 2 h,再用100 μg/ml的ox-LDL处理24 h。
1.2.2siRNA转染 根据Lipofectamine 2000转染试剂说明书,分别将Nrf2和HO-1的siRNA转染到HUVECs。转染6 h后,更换新的培养基。转染24 h后,山奈酚处理2 h,再用100 μg/ml的ox-LDL处理24 h。
1.2.3实验分组 将细胞分为6组:对照(control)组,正常培养;ox-LDL组,100 μg/ml的ox-LDL处理24 h;Kaempferol+ox-LDL组,50 μmol/L的山奈酚处理2 h后再用100 μg/ml的ox-LDL处理24 h;Kaempferol+ox-LDL+compound C组,5 mmol/L的compound C处理2 h,然后50 μmol/L的山奈酚处理2 h,再用100 μg/ml的ox-LDL处理24 h;Kaempfe-rol+ox-LDL+si-Nrf2组,si-Nrf2转染24 h,然后50 μmol/L的山奈酚处理2 h,再用100 μg/ml的ox-LDL处理24 h;Kaempferol+ox-LDL+si-HO-1组,si-HO-1转染24 h,然后50 μmol/L的山奈酚处理2 h,再用100 μg/ml的ox-LDL处理24 h。
1.2.4MTT检测细胞活力 将细胞接种到96孔板上,每组设置4个复孔,分别进行不同处理。然后每孔加入10 μl MTT试剂(0.5 mg/ml),在培养箱中孵育4 h后弃去培养基,每孔加入150 μl二甲基亚砜溶液,震荡溶解15 min,在酶标仪490 nm处检测吸光度(A)。
1.2.5流式细胞术检测细胞凋亡 将细胞接种到6孔板上,每组设置4个复孔,分别进行不同处理。然后经胰酶(不含EDTA)消化,PBS洗涤、离心后,弃上清液。每孔加入500 μl 结合缓冲液重悬细胞,然后加入10 μl annexin V-FITC,5 μl PI,避光室温孵育15 min,上流式细胞仪检测细胞凋亡。
1.2.6ROS含量的测定 将细胞接种到6孔板上,每组设置4个复孔,分别进行不同处理。然后每孔加入10 μmol/L DCFH-DA避光孵育30 min,流式细胞术分析胞间ROS水平。
1.2.7SOD含量测定 根据试剂盒说明书进行SOD含量测定。
1.2.8蛋白提取 将细胞接种到24孔板上,每组设置4个复孔,分别进行不同处理。每孔加入RIPA裂解液与PMSF的混合液(100∶1)裂解细胞15 min,离心收集上清提取总蛋白。根据试剂盒说明书提取核蛋白。BCA 蛋白试剂盒进行蛋白定量。
1.2.9Western blot检测蛋白水平 取适量样品蛋白进行SDS-PAGE凝胶电泳,采用半干转方法将蛋白质转移到硝酸纤维素膜上,然后用5%脱脂奶粉室温封闭1 h。加入Ⅰ抗4℃过夜孵育,之后加入Ⅱ抗室温孵育1 h。用Odyssey红外激光成像系统进行扫描。

2 结果
2.1山奈酚提高ox-LDL诱导的内皮细胞活性 山奈酚的化学结构如图1A所示。MTT试剂盒检测细胞活力,结果如图1B所示。与对照组相比,ox-LDL处理显著降低了HUVECs的细胞活性。当加入不同浓度山奈酚预处理后,细胞活力呈山奈酚浓度依赖性提高。当山奈酚浓度大于30 μmol/L时,细胞活力显著上升。
2.2山奈酚降低ox-LDL诱导的内皮细胞凋亡 流式细胞术检测细胞凋亡,结果如图2A和2B所示。与对照组相比,ox-LDL处理显著提高了细胞的凋亡水平。与ox-LDL处理组相比,山奈酚预处理能够呈浓度依赖性降低细胞凋亡,当浓度大于10 μmol/L时,差异具有显著统计学意义。我们进一步通过Western blot检测凋亡相关蛋白的表达,结果如图2C、D。与对照组相比,ox-LDL组的cleaved-caspase-3的蛋白水平显著上调,而Bcl-2的水平显著降低。山奈酚预处理呈浓度依赖性下调ox-LDL诱导的cleaved-caspase-3的蛋白水平,上调Bcl-2的表达。后续实验选择50 μmol/L作为山奈酚的处理浓度。
2.3山奈酚下调ox-LDL诱导的炎性因子表达 Western blot检测TNF-α、IL-1β和IL-6的蛋白表达,结果如图3所示。与对照组相比,ox-LDL处理显著上调TNF-α、IL-1β和IL-6的蛋白水平。与ox-LDL组相比,山奈酚预处理能够显著下调ox-LDL诱导的TNF-α、IL-1β和IL-6表达。
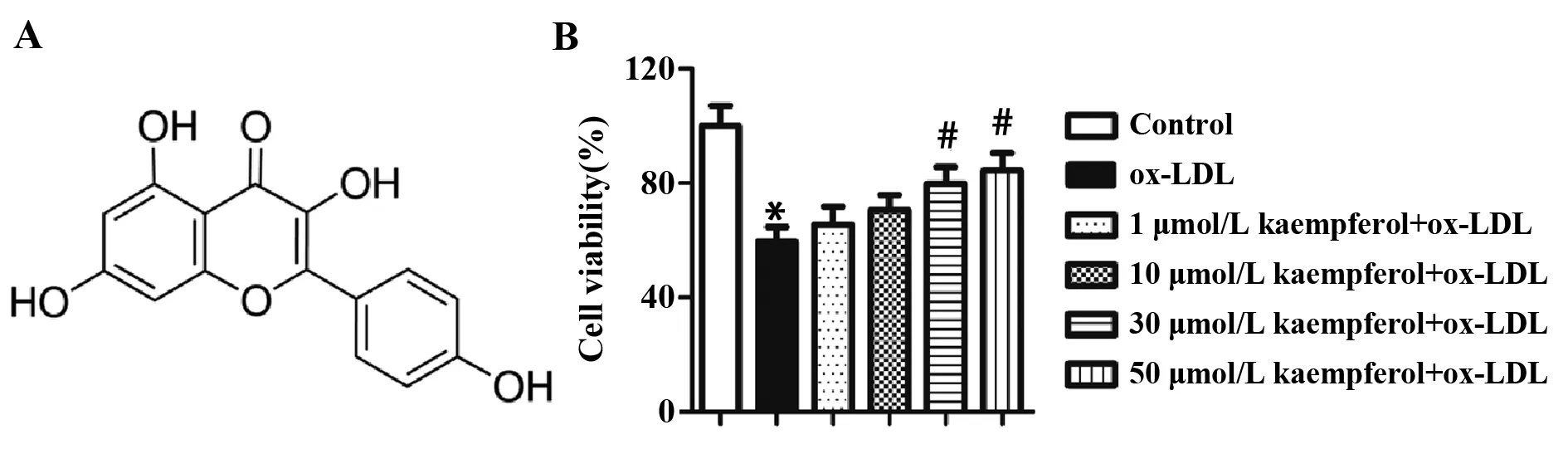
图1 山奈酚对ox-LDL诱导的HUVECs细胞活力的影响Fig.1 Effects of kaempferol on cell viability in ox-LDL-treated HUVECsNote: vs control group;#.P<0.05 vs ox-LDL group.
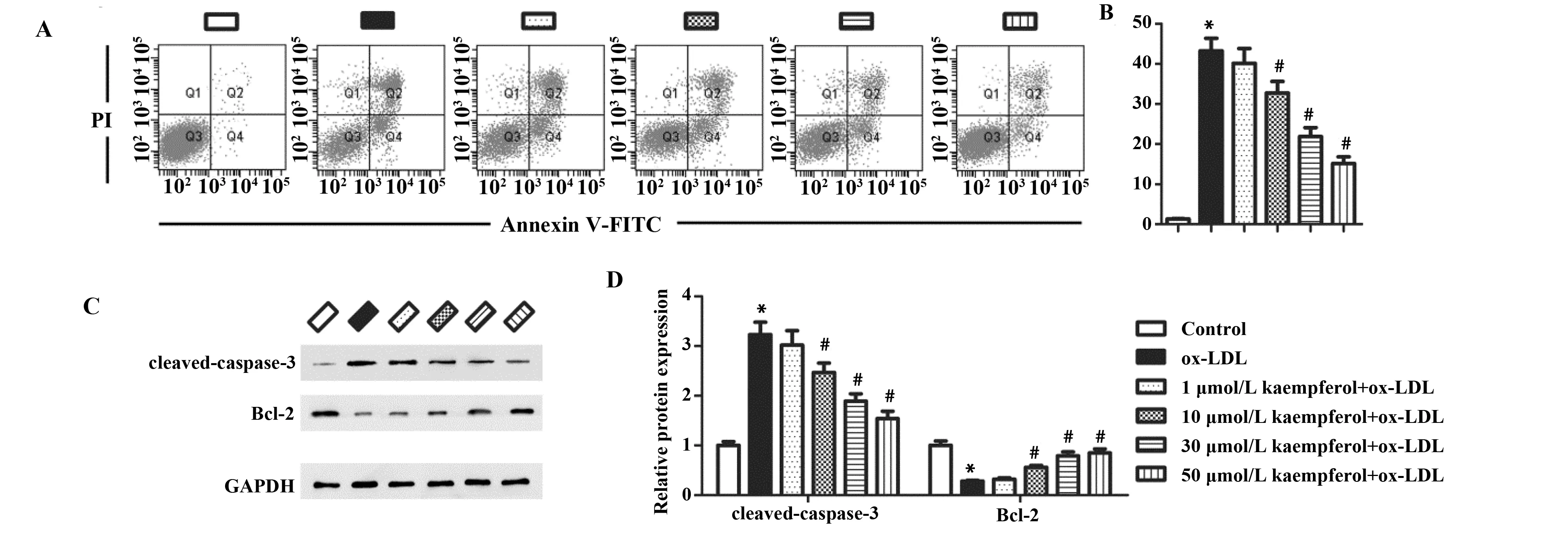
图2 山奈酚对ox-LDL诱导的HUVECs细胞凋亡的影响Fig.2 Effects of kaempferol on cell apoptosis in ox-LDL-treated HUVECsNote: vs control group;#.P<0.05 vs ox-LDL group.
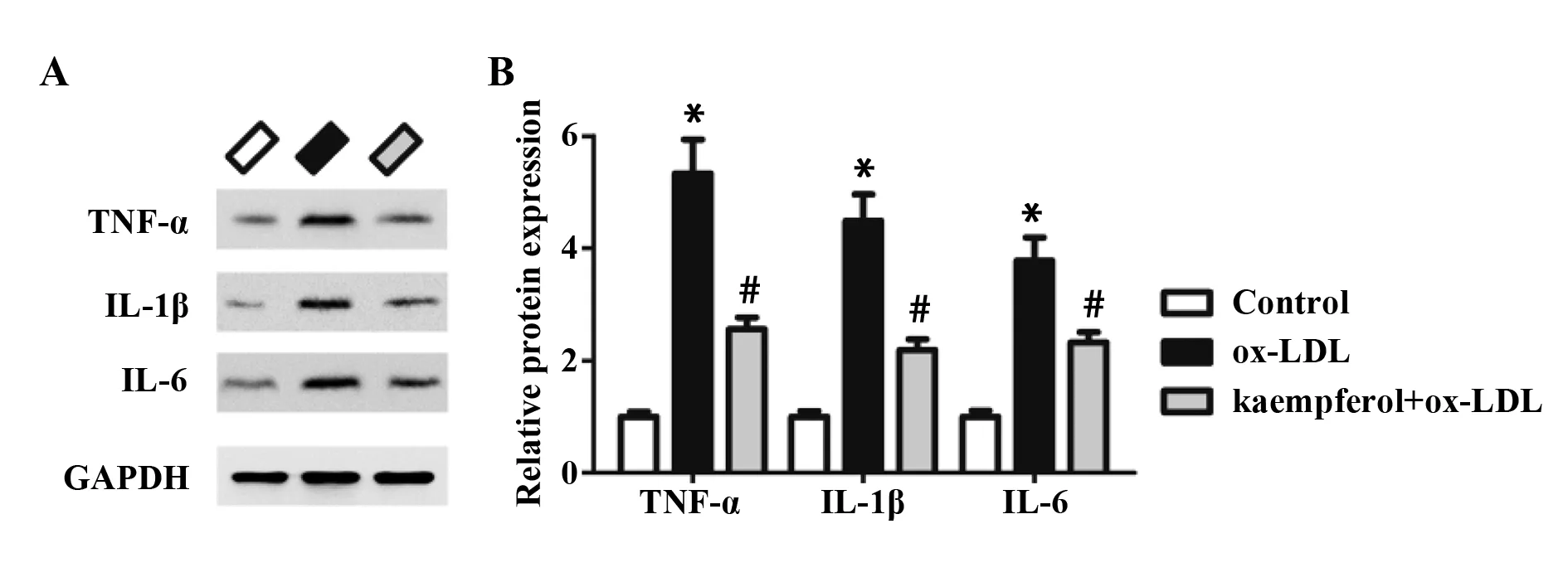
图3 山奈酚对ox-LDL诱导的HUVECs中TNF-α、IL-1β和IL-6表达的影响Fig.3 Effects of kaempferol on expression of TNF-α,IL-1β and IL-6 in ox-LDL-treated HUVECsNote: vs control group;#.P<0.05 vs ox-LDL group.
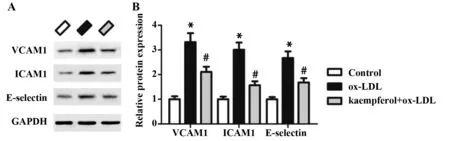
图4 山奈酚对ox-LDL诱导的HUVECs中VCAM1、ICAM1和E-selectin表达的影响Fig.4 Effects of kaempferol on expression of VCAM1,ICAM1 and E-selectin in ox-LDL-treated HUVECsNote: vs control group;#.P<0.05 vs ox-LDLgroup.
2.4山奈酚下调ox-LDL诱导的黏附分子表达 Western blot检测血管细胞黏附分子(VCAM1)、细胞间黏附分子(ICAM1)和选择素E(E-selectin)的蛋白表达,结果如图4所示。与对照组相比,VCAM1、ICAM1和E-selectin的蛋白表达水平在ox-LDL组中显著上调,而山奈酚预处理能够显著下调VCAM1、ICAM1和E-selectin的表达。
2.5山奈酚抑制ox-LDL诱导的氧化应激 如图5所示,与对照组相比,ox-LDL处理能够显著上调ROS的产生,下调SOD的活性。与ox-LDL组相比,山奈酚预处理能够显著下调ox-LDL诱导的ROS产生,上调SOD 的活性。
2.6山奈酚通过激活AMPK-Nrf2-HO-1通路抑制氧化应激损伤 Western blot检测p-AMPK、AMPK、Nrf2和HO-1的蛋白表达,结果如图6A~C所示。与对照组相比,p-AMPK、Nrf2和HO-1在ox-LDL组中的蛋白水平显著下降,而山奈酚预处理显著提高p-AMPK、Nrf2和HO-1的蛋白水平。为了进一步确定AMPK-Nrf2-HO-1通路与山奈酚的保护作用紧密相关,我们用AMPK抑制剂compound C、si-Nrf2和si-HO-1进行处理来抑制该通路蛋白的表达。如图6A~C所示,与kaempfe-rol+ox-LDL组相比,C.C+kaempferol+ox-LDL组中p-AMPK、Nrf2和HO-1的蛋白水平均显著下调,si-Nrf2+kaempferol+ox-LDL组中Nrf2和HO-1的蛋白水平显著下调,p-AMPK表达无显著变化,而si-HO-1+kaempferol+ox-LDL组中HO-1的蛋白水平显著下调,p-AMPK和Nrf2表达无显著变化。这个结果说明AMPK通过调控Nrf2影响HO-1的表达。与kaempferol+ox-LDL组相比,Compound C、si-Nrf2和si-HO-1处理组都能够增加ROS的生成(图6D)。进一步通过MTT实验和流式细胞术检测发现,与kaempferol+ox-LDL组相比,Compound C、si-Nrf2和si-HO-1处理组的细胞活力显著下降,而细胞凋亡显著增加(图6E和6F)。这些结果说明山奈酚能够通过激活AMPK-Nrf2-HO-1通路降低ox-LDL诱导的氧化应激等损伤。
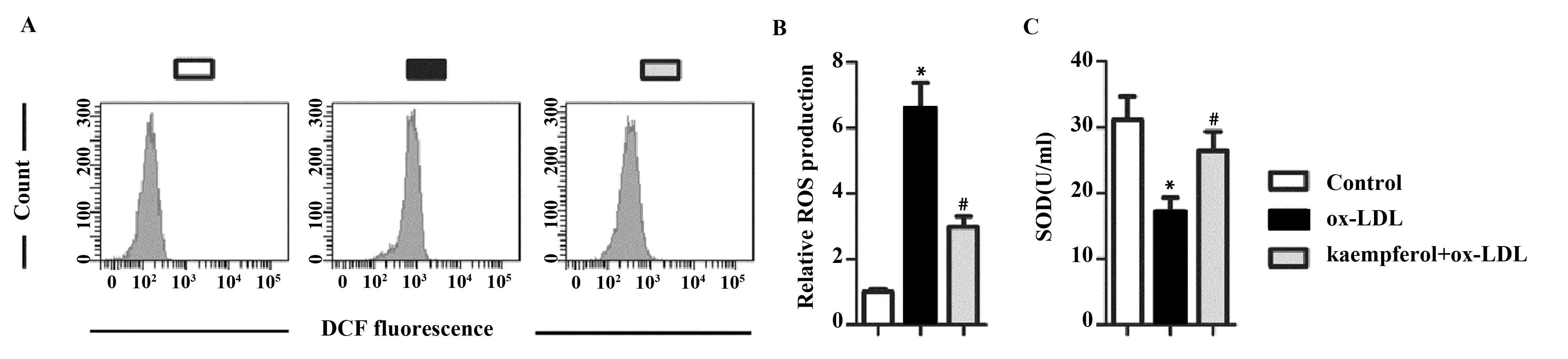
图5 Effects of kaempferol on oxidative stress in ox-LDL-treated HUVECsFig.5 山奈酚对ox-LDL诱导的HUVECs中氧化应激的影响Note: vs control group;#.P<0.05 vs ox-LDL group.
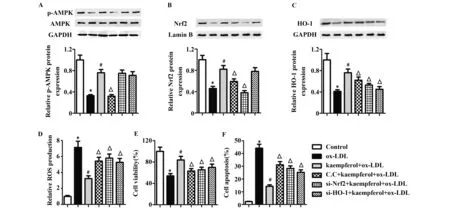
图6 山奈酚对ox-LDL诱导的HUVECs中AMPK-Nrf2-HO-1通路的影响Fig.6 Effects of kaempferol on AMPK-Nrf2-HO-1 signaling in ox-LDL-treated HUVECsNote: vs control group;#.P<0.05 vs ox-LDL group;△.P<0.05 vs kaempferol+ox-LDL group.
3 讨论
AS是一种常见的心血管疾病,可由高血压、高血糖、高血脂、吸烟、饮酒等多种因素引发,严重威胁人类健康。血管内皮损伤是AS的主要病理基础,是一个包括凋亡、炎症和氧化应激的复杂过程。本研究发现山奈酚能够缓解ox-LDL诱导的HUVECs损伤,并初步探讨了其分子机制,发现山奈酚能够通过激活AMPK/Nrf2/HO-1信号通路降低氧化应激反应,从而减少内皮损伤。
在正常生理状态下,内皮细胞的增殖和凋亡保持动态平衡,以此来维持血管的正常功能。研究表明,在AS斑块中存在广泛的内皮细胞凋亡,而内皮细胞的过度凋亡是内皮功能障碍的始动环节。在本研究中,我们首先通过MTT实验检测细胞活力,发现ox-LDL处理显著降低了HUVECs的细胞活力。当用山奈酚预处理后,HUVECs的细胞活力呈浓度依赖性提高,说明山奈酚能够缓解ox-LDL诱导的细胞活力损伤。流式细胞术检测细胞凋亡,发现ox-LDL处理显著提高HUVECs的凋亡率,而山奈酚能够呈浓度依赖性降低ox-LDL诱导的细胞凋亡。我们进一步通过Western blot检测了凋亡相关蛋白的表达,发现山奈酚能够呈浓度依赖性下调ox-LDL诱导的cleaved-caspase-3的蛋白水平,上调Bcl-2的表达。以上结果说明山奈酚能够保护HUVECs抵抗ox-LDL诱导的损伤,且保护作用呈浓度依赖性。
血管内皮的炎性反应在AS的发生发展过程中发挥重要的作用,白细胞向血管内皮的滚动、黏附、迁移和积聚,是血管内皮炎性反应的标志[10]。内皮细胞在外界环境刺激下,能够诱导多种炎性因子和黏附分子的产生,加速白细胞向内皮细胞的黏附和炎性反应。Jiang等[11]研究显示,ox-LDL处理HUVECs后,IL-6、TNF-α、VCAM1、ICAM1和E-selectin的含量明显升高。有文献指出,山奈酚能够通过降低炎性因子和黏附因子的表达发挥抗AS作用[12,13]。我们的研究结果与以上结果一致,ox-LDL处理显著上调了TNF-α、IL-1β、IL-6、VCAM1、ICAM1和E-selectin的表达,而山奈酚预处理使以上蛋白的表达显著下调。以上结果说明山奈酚能够保护HUVECs抵抗ox-LDL诱导的炎性损伤。
有文献报道,细胞增殖凋亡异常以及炎性反应与氧化应激有密不可分的关系。当ROS的生物活性高于抗氧化防御能力时,即产生氧化应激反应。在多种心血管疾病中,例如AS代谢综合征、血脂异常,ROS水平明显升高,与疾病的发生发展紧密相关[14,15]。SOD是主要的抗氧化酶,能够保护机体免受氧化损伤[16]。本研究结果发现,ox-LDL处理能够显著上调ROS的产生,下调SOD的表达。山奈酚预处理后能够显著下调ox-LDL诱导的ROS产生,上调SOD 的表达,与Xiao等[8]的研究结果一致。这个结果说明山奈酚能够保护HUVECs抵抗ox-LDL诱导的氧化应激损伤。
Nrf2 是一个调控多种抗氧化酶基因表达的重要的转录因子,例如HO-1[17,18],一种具有抗氧化和细胞保护作用的酶[19,20]。Nrf2/HO-1 信号的激活能够保护细胞免受氧化损伤。AMPK是一种丝/苏氨酸蛋白激酶,有研究报道AMPK的活化能够抑制异常炎症反应和氧化应激[21]。本实验的Western blot检测发现,ox-LDL处理显著下降p-AMPK、Nrf2和HO-1的蛋白水平,而山奈酚预处理显著提高了p-AMPK、Nrf2和HO-1的蛋白水平。山奈酚能够通过提高Nrf2的表达来降低ROS的产生[22],AMPK在癌症中起到调控作用[23],为了进一步确定AMPK-Nrf2-HO-1通路与山奈酚的保护作用紧密相关,我们用AMPK抑制剂compound C、si-Nrf2和si-HO-1进行处理来抑制该通路蛋白的表达。与kaempferol+ox-LDL组相比,Compound C、si-Nrf2和si-HO-1处理组都能够提高ROS的生成。进一步通过MTT实验和流式细胞术检测发现,与kaempferol+ox-LDL组相比,Compound C、si-Nrf2和si-HO-1处理组的细胞活力显著下降,而细胞凋亡显著增加。这些结果说明山奈酚能够通过激活AMPK-Nrf2-HO-1通路降低ox-LDL诱导的氧化应激等损伤。
综上所述,山奈酚抑制ox-LDL诱导的HUVECs活力降低,减少细胞凋亡,下调炎性因子和黏附分子的表达,缓解氧化应激反应,这与AMPK-Nrf2-HO-1信号通路的激活有关。本研究进一步阐明了内皮损伤及AS的发病机理,为山奈酚防治AS提供了理论依据。
参考文献:
[1] Gimbrone MA,García-Cardea G.Endothelial cell dysfunction and the pathobiology of atherosclerosis[J].Circ Res,2016,118(4):620-636.
[2] 姚树桐,李严严,刘庆华,等.氧化低密度脂蛋白通过 CD36介导的氧化应激诱导巨噬细胞自噬[J].中国病理生理杂志,2015,31(6):1002-1007.
Yao ST,Li YY,Liu QH,etal.Oxidized low-density lipoprotein induces autophagy in macrophages via CD36-mediated oxidative stress[J].Chin J Pathophy,2015,31( 6):1002-1007.
[3] 李严严,徐晓燕,张家君,等.蜂胶醇提物通过抑制caspase-12减轻氧化低密度脂蛋白诱导的巨噬细胞凋亡[J].中国病理生理杂志,2015,31(12):2202-2208.
Li YY,Xu XY,Zhang JJ,etal.Ethanol extract of propolis protects macrophages from oxidized low-density lipoprotein-induced apoptosis by inhibiting caspase-12[J].Chin J Pathophy,2015,31(12):2202-2208.
[4] 肖坤福,廖晓峰,于 容,等.黄酮类化合物保护低密度脂蛋白氧化作用和减少动脉粥样硬化作用[J].广州食品工业科技,2004,20(1):100-103.
Xiao KF,Liao XF,Yu R,etal.Effects of Flavonoids protecting against low-density lipoprotein oxidation and atherosclerosis[J].Guangzhou Food Science and Technology,2004,20(1):100-103.
[5] Chen AY,Chen YC.A review of the dietary flavonoid,kaempferol on human health and cancer chemoprevention[J].Food Chem,2013,138(4):2099-2107.
[6] Rajendran P,Rengarajan T,Nandakumar N,etal.Kaempferol,a potential cytostatic and cure for inflammatory disorders[J].Eur J Med Chem,2014,86(51):103-112.
[7] 周明杰,刘立群.山奈酚预处理对大鼠心脏缺血再灌注损伤的影响[J].潍坊医学院学报,2015,37 (3):168-170.
Zhou MJ,Liu LQ.Effects of Kaempferol against myocardial ischemia-reperfusion injury in rat heart[J].Acta Acad Med Weifang,2015,37 (3):168-170.
[8] Xiao HB,Fang J,Lu XY,etal.Protective effects of kaempferol against endothelial damage by an improvement in nitric oxide production and a decrease in asymmetric dimethylarginine level[J].Eur J Pharm,2009,616(1-3):213-222.
[9] Salvamani S,Gunasekaran B,Shaharuddin NA,etal.Antiartherosclerotic effects of plant flavonoids[J].Biomed Res Int,2014,2014(1):480258.
[10] 潘玉婷,郭春雨,马晓娟,等.氧化低密度脂蛋白损伤血管内皮细胞的机制及雌激素保护作用[J].中华老年心脑血管病杂志,2014,16(2):209-212.
Pan YT,Guo CY,Ma XJ,etal.Mechanism of oxidized low density lipoprotein on vascular endothelial cells and protective effect of estrogen[J].Clin J Geriatr Heart Brain Vessel Dis,2014,16(2):209-212.
[11] Jiang YR,Miao Y,Yang L,etal.Effect of chinese herbal drug-containing serum for activating-blood and dispelling-toxin on ox-LDL-induced inflammatory factors′expression in endothelial cells[J].Chin J Integrative Med,2012,18(1):30-33.
[12] Crespo I,García-Mediavilla MV,Gutiérrez B,etal.A comparison of the effects of kaempferol and quercetin on cytokine-induced pro-inflammatory status of cultured human endothelial cells[J].Bri J Nut,2008,100(5):968-976.
[13] Kong L,Luo C,Li X,etal.The anti-inflammatory effect of kaempferol on early atherosclerosis in high cholesterol fed rabbits[J].Lipids Health Dis,2013,12:115.
[14] Ilkun O,Boudina S.Cardiac dysfunction and oxidative stress in the metabolic syndrome:an update on antioxidant therapies[J].Curr Pharm Design,2013,19(27):4806-4817.
[16] Cho IH,Gong JH,Kang MK,etal.Astragalin inhibits airway eotaxin-1 induction and epithelial apoptosis through modulating oxidative stress-responsive MAPK signaling[J].BMC Pulmonary Med,2014,14(1):1-11.
[17] Alam J,Cook JL.Transcriptional regulation of the heme oxygenase-1 gene via the stress response element pathway [J].Curr Pharm Design,2003,9(30):2499-2511.
[18] Chen XL,Dodd G,Thomas S,etal.Activation of Nrf2/ARE pathway protects endothelial cells from oxidant injury and inhibits inflammatory gene expression[J].Am J Physiol Heart Circ Physiol,2006,290(5):H1862-H1870.
[19] Yuan F,Tu S,Qiu X,etal.Protective effect of alpha-mangostin against oxidative stress induced-retinal cell death[J].Sci Reports,2016,6:21018.
[20] Lawal AO,Marnewick JL,Ellis EM.Heme oxygenase-1 attenuates cadmium-induced mitochondrial-caspase 3-dependent apoptosis in human hepatoma cell line[J].BMC Pharmacol Toxicol,2015,16(1):1-13.
[21] Huang BP,Lin CH,Chen HM,etal.AMPK activation inhibits expression of proinflammatory mediators through downregulation of PI3K/p38 MAPK and NF-κB signaling in murine macrophages[J].DNA Cell Biol,2015,34(2):133-141.
[22] Saw CL,Guo Y,Yang AY,etal.The berry constituents quercetin,kaempferol,and pterostilbene synergistically attenuate reactive oxygen species:involvement of the Nrf2-ARE signaling pathway[J].Food Chem Toxicol,2014,72(8):303-311.
[23] 李君茹,李会方,周 嘉,等.二甲双胍对U937细胞增殖、周期及凋亡的影响[J].中国免疫学杂志,2017,33(9):1315-1319.
Li JR,Li HF,Zhou J,etal.Effect of metformin on proliferation,cell cycle and apoptosis of U937 cells[J].Chin J Immunol,2017,33(9):1315-1319.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
温州大学学报(自然科学版)(2022年2期)2022-05-30
昆明医科大学学报(2022年4期)2022-05-23
上海交通大学学报(医学版)(2022年3期)2022-05-05
现代临床医学(2021年5期)2021-11-02
潍坊学院学报(2020年2期)2021-01-18
世界科学技术-中医药现代化(2020年2期)2020-07-25
中国眼镜科技杂志(2019年9期)2019-11-11
制导与引信(2017年3期)2017-11-02
中国当代医药(2015年30期)2015-03-01
