
姜黄素对宫内发育迟缓断奶仔猪肠道抗氧化功能的影响
2019-08-30 06:12何进田沈明明张莉莉
食品科学 2019年15期
王 斐,何进田,沈明明,张 昊,牛 玉,张莉莉,王 恬*
(南京农业大学动物科技学院,江苏 南京 210095)
姜黄素是从姜科姜黄属(Curcumin longa)植物的根茎中提取的一种天然酚类化合物,广泛用于香料和食品添加剂[1]。研究发现,姜黄素具有抗氧化、抗炎、降糖、降压、降血脂、保护黏膜和免疫调节等生理作用[2]。作为一种天然抗氧化物质,姜黄素可通过清除自由基、提高抗氧化酶活力、调节谷胱甘肽(glutathione,GSH)代谢效率、激活核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)信号途径等方式,改善氧化还原平衡,以发挥细胞保护作用[3-6]。随着研究的不断深入,姜黄素的抗氧化作用引起了业内学者的广泛关注。2014年,姜黄素被列入饲料添加剂目录[7],其作为一种绿色、安全的新型添加剂也成为了国内外的研究热点之一。
前人研究表明,姜黄素可减少氧化应激状态下小鼠肠道多种解毒酶的异常表达,如谷胱甘肽硫转移酶(glutathione S-transferase,GST)、NAD(P)H:醌氧化还原酶1(NAD(P)H : quinone oxidoreductase l,NQO1)以及血红素加氧酶1(heme oygenase 1,HO-1)[1-2]。荀文娟等[8]研究结果显示,添加400 mg/kg姜黄素可改善断奶仔猪回肠黏膜的形态结构,增强肠道黏膜屏障的完整性。此外,在日粮中添加400 mg/kg姜黄素可显著提高大肠杆菌攻毒仔猪的生长性能,修复肠道黏膜紧密连接,提高肠道抗氧化与免疫功能[9]。
宫内发育迟缓(intrauterine growth retardation,IUGR)是围产期常见的并发症之一,是指哺乳动物的胚胎或胎儿在子宫内生长发育受阻的现象。IUGR不仅会导致胎儿在新生期的发病率与死亡率升高,对其出生后较长时期的生长发育与健康状况也会造成不可忽视的负面影响[10-11]。已有研究表明,IUGR新生动物的肠道损伤明显,表现为绒毛形态受损、消化吸收能力较差、抗氧化系统发育不完善等[9,12-13]。目前,针对IUGR动物肠道损伤的营养调控措施较为缺乏,关于姜黄素类抗氧化剂的应用效果更是鲜见报道。鉴于姜黄素良好的抗氧化性及其在保护动物肠道功能上的积极作用,本实验以IUGR断奶仔猪为研究对象,系统地探讨了姜黄素对其肠道抗氧化功能的影响。同时,由于猪在解剖学、生理学以及代谢特点等方面均与人类有较高的相似性[14-15],因此本研究结果也可为IUGR新生儿肠道疾病的预防与治疗提供新的思路与参考依据。
1 材料与方法
1.1 动物、材料与试剂
实验仔猪(杜洛克×长白×大白)选自江苏立华牧业股份有限公司宿迁分公司养殖场。本实验经南京农业大学动物护理和使用委员会批准,许可证号:NJAUCAST-2015-098。
姜黄素(纯度≥98%) 广州科虎生物技术研究开发中心;丙二醛(malondialdehyde,MDA)试剂盒、蛋白质羰基(protein carbonyl,PC)试剂盒、总超氧化物歧化酶(total superoxide dismutase,T-SOD)试剂盒、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)试剂盒、GSH试剂盒、过氧化氢酶(catalase,CAT)试剂盒、总抗氧化能力(total antioxidant capacity,T-AOC)试剂盒、过氧化氢(H2O2)试剂盒、总蛋白定量测定(BCA法)试剂盒 南京建成生物工程研究所;8-羟脱氧鸟苷(8-hydroxy-2'-deoxyguanosine,8-OHDG)试剂盒上海易利生物科技有限公司;TRIzol试剂 美国英杰生命技术有限公司;mRNA反转录试剂盒、SYBR实时聚合酶链式反应(quantitative real-time polymerase chain reaction,qPCR)试剂盒 日本TaKaRa生物科技股份有限公司。
1.2 仪器与设备
BSA224S-CW型分析天平 德国Sartorius公司;Bio-Gen Series PRO200型匀浆机 美国PRO Scientific公司;5804R型台式高速冷冻离心机 德国Eppendorf公司;DK-S24型电热恒温水浴锅 上海精宏实验设备有限公司;T-6m型紫外-可见分光光度计 南京菲勒仪器有限公司;DW-25L262型医用低温保存箱 青岛海尔特种电器有限公司;Multiskan Go型全波长酶标仪、8925型超低温冰箱(-80 ℃)、实时荧光PCR仪 美国Thermo Fisher Scientific公司。
1.3 方法
1.3.1 实验动物和实验设计
动物实验设计及程序按照《中国实验动物管理条例》的要求。选取胎次、体质量接近以及遗传基础一致的妊娠母猪20 头,在其分娩当日,从每窝新生仔猪中选择1 头正常初生体质量(normal birth mass,NBM)和1 头IUGR仔猪。以初生体质量低于群体2 个标准差的仔猪为IUGR仔猪,而初生体质量在群体1 个标准差范围内的仔猪为NBM仔猪[16]。本实验中以初生体质量(1.51±0.04)kg为NBM仔猪,初生体质量低于(0.96±0.02)kg为IUGR仔猪。26 日龄断奶时,将NBM仔猪随机均分为N组和NC组,IUGR仔猪随机均分为I组和IC组(每组10 头,公母各半)。N组和I组饲喂基础日粮,NC和IC组在基础日粮上添加400 mg/kg姜黄素(即每千克基础日粮添加400 mg姜黄素),饲喂至50 日龄。
1.3.2 饲养管理
实验期内仔猪每组饲养于一栏,每头猪平均占有圈舍1 m×0.6 m,自由采食和饮水,每天清扫和消毒猪舍,猪舍温度和相对湿度分别控制在25~28 ℃和50%~70%。按猪场饲养管理要求进行驱虫和常规免疫,并观察和记录动物的健康状况。日粮组成及水平参照美国国家研究委员会(National Research Council,NRC)(2012)推荐营养水平配制,其组成及营养成分见表1。
有次去,剃头师傅不在,店铺里空着,没有人,就往后面的院子里走,边走边问可有人,这时出来一个胖胖的妇人,问我找谁。

表1 日粮配方和营养水平Table 1 Composition and nutrient levels of diets
1.3.3 样品采集
仔猪饲养至50 日龄时,分别从每组选择8 头,宰前空腹12 h,将仔猪电击致晕,放血致死。而后迅速剖开腹腔,取出肠道并分离出空肠和回肠。在各段肠道中部分别取一段15 cm的组织,纵行剪开后用磷酸盐缓冲液(phosphate buffered saline,PBS)冲洗干净后置于洁净滤纸上,用灭菌载玻片刮取肠黏膜并装入冻存管于液氮中速冻,于-80 ℃保存以备后期指标测定。
1.3.4 指标测定
1.3.4.1 抗氧化性能检测
采用BCA法测定蛋白浓度,严格按照产品说明书进行操作。肠道黏膜中MDA、PC、T-SOD、T-AOC、CAT、GSH-Px、GSH和H2O2水平均采用南京建成生物工程研究所试剂盒进行检测,并严格按照产品说明书要求进行操作。
1.3.4.2 肠道黏膜中的细胞因子检测
采用酶联免疫吸附测定试剂盒检测肠道黏膜中8-OHDG含量,测定过程严格按照试剂盒说明书的方法和步骤进行。
采用qPCR检测肠道黏膜相关基因mRNA表达量。取组织样品200~300 mg,采用TRIzol试剂提取总RNA。检测所有样品RNA质量浓度和提取完整性,并用体积分数0.1%的焦碳酸二乙酯将各样品RNA质量浓度调整至500 ng/μL。采用相同质量浓度的总RNA样品按照反转录试剂盒说明书,加入总RNA 2.0 μL、20×RT Enzyme Mix 4.0 μL、Nuclease-free H2O 14.0 μL,按照下列条件进行反转录反应:37 ℃,15 min;85 ℃,5 s;4 ℃保存。反应结束后所得的cDNA原液用超纯水按体积比1∶4稀释。按照SYBR®Premix ExTaq™定量试剂盒说明书进行操作,依次往qPCR管中加入以下试剂(20 μL体系):cDNA 2 μL、SYBR Premix ExTaq10 μL、上游引物0.4 μL、下游引物0.4 μL、ROX Reference Dye 0.4 μL、超纯水6.8 μL,混匀离心后在qPCR仪上反应检测。反应程序如下:95 ℃、0 min;95 ℃、15 s,60 ℃、45 s,重复40 个循环。选择β-actin作为内参基因,利用2-ΔΔCt法对目的基因mRNA表达进行相对定量。各目的基因引物序列见表2。

表2 qPCR引物序列Table 2 Primer sequences used for real-time PCR
1.4 数据统计与分析
数据经Excel 2016软件初步整理后,使用SPSS 20.0软件进行统计分析,结果以平均值±标准差表示。各处理组间差异采用GLM模块的双因素方差分析,并用Tukey法进行多重比较,以仔猪初生体质量(B)和日粮添加姜黄素(400 mg/kg)(D)作为实验因子,并检验两因子之间的互作效应。P<0.05作为差异显著性判断标准。
2 结果与分析
2.1 姜黄素对IUGR断奶仔猪肠道MDA、PC和8-OHDG含量的影响
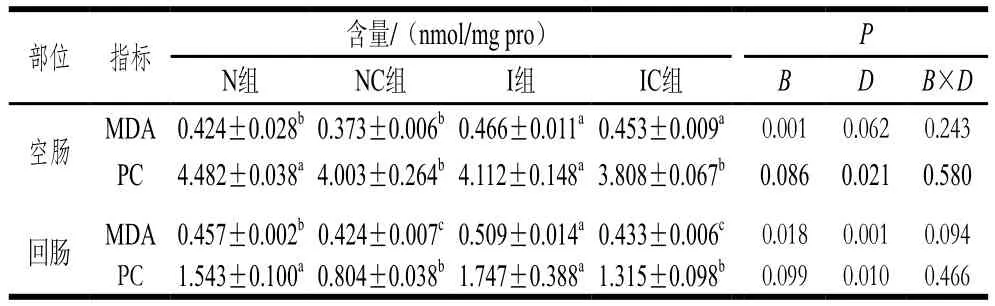
表3 姜黄素对IUGR断奶仔猪肠道中MDA和PC含量的影响Table 3 Effect of curcumin on the contents of MDA and PC in intestinal tissues of weaning piglets with IUGR

表4 姜黄素对IUGR断奶仔猪肠道中8-OHDG含量的影响Table 4 Effect of curcumin on 8-OHDG content in intestinal tissues of weaning piglets with IUGR
由表3、4可知,对于断奶仔猪空肠,IUGR仔猪的MDA、8-OHDG含量升高;日粮中添加姜黄素后,IC组PC、8-OHDG含量显著降低(P<0.05),但对MDA含量无显著影响(P>0.05);初生体质量和日粮添加姜黄素对空肠8-OHDG含量具有显著的互作效应(P<0.05)。对于断奶仔猪的回肠部位,与N组相比,I组MDA含量显著升高(P<0.05),PC、8-OHDG含量无显著差异(P>0.05);NC组MDA、PC含量显著降低(P<0.05),8-OHDG含量无显著差异(P>0.05);与I组相比,IC组MDA、PC含量显著降低(P<0.05),8-OHDG含量无显著差异(P>0.05)。
2.2 姜黄素对IUGR断奶仔猪肠道T-AOC、抗氧化酶活力、GSH和H2O2含量的影响
由表5可知,与N组相比,I组断奶仔猪空肠中T-SOD活力和GSH含量显著降低(P<0.05),T-AOC降低29.2%(P>0.05),GSH-Px活力降低34.4%(P>0.05);日粮中添加姜黄素后,与N组和I组相比,NC组和IC组的T-AOC、CAT活力显著升高(P<0.05),GSH含量显著升高(P<0.05),H2O2含量分别降低14.4%、5.8%(P>0.05);初生体质量和日粮添加姜黄素对断奶仔猪空肠GSH含量和GSH-Px活力有显著的互作效应(P<0.05)。

表5 姜黄素对IUGR断奶仔猪空肠T-AOC、抗氧化酶活力、GSH和H2O2含量的影响Table 5 Effect of curcumin on T-AOC, antioxidant enzymes activities and GSH and H2O2content in the jejunum of weaning piglets with IUGR

表6 姜黄素对IUGR断奶仔猪回肠T-AOC、抗氧化酶活力、GSH含量和H2O2含量的影响Table 6 Effect of curcumin on T-AOC, antioxidant enzymes activities and GSH and H2O2contents in the ileum of weaning piglets with IUGR
由表6可知,断奶仔猪回肠中,I组T-SOD活力、GSH含量较N组分别显著降低13.9%、18.1%(P<0.05),I组H2O2含量较N组显著升高14.3%(P<0.05)。与N组和I组相比,日粮中添加姜黄素后,NC组和IC组中H2O2含量均显著降低(P<0.05);与N组相比,NC组T-AOC升高10.4%(P>0.05);与I组相比,IC组CAT活力显著升高(P<0.05),T-AOC和GSH-Px活力分别升高19.1%和37.4%(P>0.05)。初生体质量和日粮添加姜黄素对断奶仔猪回肠CAT和GSH-Px活力有显著的互作效应(P<0.05)。
2.3 姜黄素对IUGR断奶仔猪肠道抗氧化基因mRNA表达量的影响
由表7可知,与N组相比,I U G R仔猪空肠中Nrf2、KEAP1、SOD、GSH-Px1、GST、NQO1、GCLM、HO-1、TXNRD1的mRNA表达水平显著降低(P<0.05),CAT的mRNA表达量下降20.7%(P>0.05)。日粮中添加姜黄素显著升高了Nrf2、CAT、SOD、GST、NQO1的mRNA表达水平(P<0.05)。初生体质量和日粮添加姜黄素对断奶仔猪空肠SOD和TXNRD1的mRNA表达量有显著的互作效应(P<0.05)。

表7 姜黄素对IUGR断奶仔猪空肠抗氧化基因mRNA表达量的影响Table 7 Effect of curcumin on mRNA expression antioxidant enzymes in the jejunum of weaning piglets with IUGR
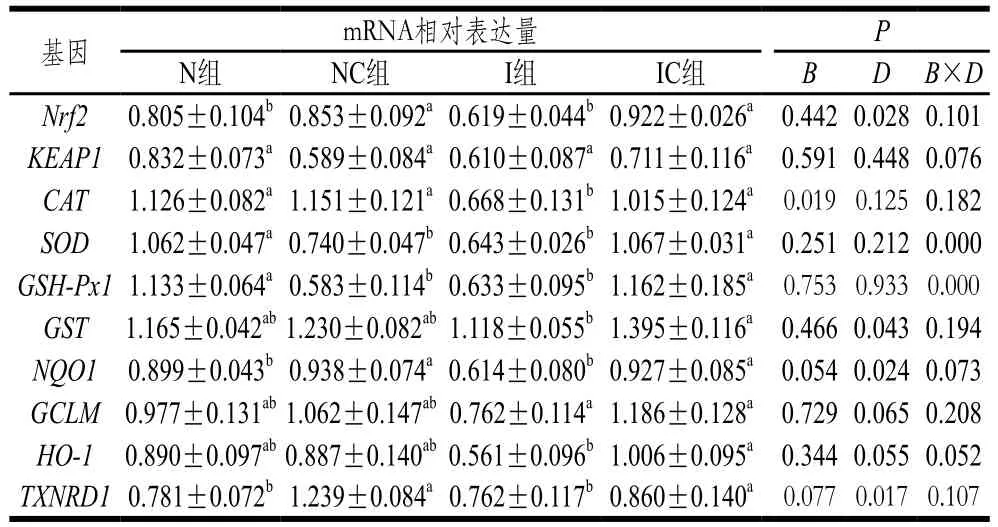
表8 姜黄素对IUGR断奶仔猪回肠抗氧化基因mRNA表达量的影响Table 8 Effect of curcumin on mRNA expression of antioxidant enzymes in the ileum of weaning piglets with IUGR
由表8可知,与N组相比,I组断奶仔猪回肠中CAT、SOD和GSH-Px1的mRNA表达量显著降低(P<0.05)。与N组相比,NC组TXNRD1、Nrf2和NQO1的mRNA表达量显著升高(P<0.05);GCLM的mRNA表达量升高8.0%(P>0.05)。与I组相比,IC组HO-1、CAT、SOD、Nrf2、GST、NQO1、TXNRD1的mRNA表达量显著升高(P<0.05);GCLM的mRNA表达量升高35.8%(P>0.05)。初生体质量和日粮添加姜黄素对断奶仔猪回肠SOD和GSH-Px1的mRNA表达量有极显著的互作效应(P<0.01)。
3 讨 论
研究表明,IUGR会使肠道发生氧化应激,对肠道造成损伤,是导致IUGR肠道损伤的重要因素之一[10]。机体中代谢和信号传递会产生少量氧自由基,同时机体的清除系统维持着氧自由基的平衡。一旦机体产生过多的氧自由基得不到及时清除就会打破这种动态平衡,引起机体发生氧化应激,产生大量氧化物,例如MDA、PC等[17]。随着社会的进步和人们营养观念的提高,通过营养干预来改善IUGR健康状况的技术愈来愈受关注。而姜黄素作为一种天然抗氧化剂,在防止或缓解机体氧化应激方面起到重要调节作用。
Takagi等[18]研究发现,8-OHDG是氧化应激引起DNA损伤所产生的脱氧鸟苷的羟基产物,IUGR中8-OHDG水平较正常组显著升高。MDA是细胞脂质过氧化反应的主要产物和生物标志物[19]。苏伟鹏等[12]研究表明,IUGR显著增加了空肠中MDA的含量,证明其脂质过氧化程度会升高。Zhang Hao等[20]研究结果也表明IUGR增加了MDA含量。本实验中,IUGR断奶仔猪空肠和回肠的MDA含量、空肠8-OHDG含量均高于正常组,这些结果提示IUGR断奶仔猪肠道可能发生了氧化应激,与前人研究相似。日粮中添加姜黄素后,NC、IC组空肠和回肠PC含量、IC组空肠8-OHDG含量均显著降低,NC组和IC组MDA含量均有降低的趋势。在研究姜黄素对大鼠肠道抗氧化性能方面,姜黄素有降低PC含量的趋势[17]。Inano等[21]研究表明,姜黄素可以改善氧化应激引起的大鼠抗氧化能力下降,以及降低8-OHDG水平。另有研究结果也显示,姜黄素能够显著降低脂多糖(lipopolysaccharide,LPS)注射后断奶仔猪空肠和回肠黏膜的MDA含量[22],与本实验中姜黄素对肠道MDA含量的降低作用相似。因此,姜黄素可能具有缓解IUGR断奶仔猪肠道氧化损伤的功能。
T-AOC代表了一个体系内各种抗氧化分子和酶的总体水平,是机体抗氧化应激损伤的重要指标[23]。T-SOD、GSH-Px和CAT均属于抗氧化酶,T-SOD可以清除活性氧(reactive oxygen species,ROS),GSHPx可以清除有机过氧化物和H2O2,CAT可以清除过H2O2
[24]。GSH在GSH-Px作用下可以清除细胞内的H2O2,在机体ROS代谢平衡方面起到调节作用[25-26]。H2O2是一种高毒性氧化剂,在细胞中不断产生,必须通过抗氧化防御酶快速解毒,H2O2含量过高会诱导细胞氧化应激产生损伤[27]。本研究发现,IUGR断奶仔猪空肠、回肠中T-SOD活力和GSH含量显著降低,H2O2含量显著升高。由此可见,IUGR可能会导致断奶仔猪肠道抗氧化酶活力和抗氧化物质含量的下降,加剧氧化应激程度。王远孝等[28]研究结果显示,IUGR仔猪肠道发生氧化应激,空肠黏膜SOD活力显著降低。He Qinghua等[29]研究发现,IUGR仔猪与正常组相比肠道中GSH含量显著下降。Huang Qiang等[30]在研究亮氨酸对断奶早期IUGR仔猪空肠氧化还原状态的保护作用时发现,空肠CAT和GSH-Px的活力并无显著影响。研究表明,IUGR仔猪空肠黏膜H2O2含量显著升高[13],这与本实验结果相似。这些结果均表明IUGR仔猪的抗氧化能力会出现一定程度的下降。本实验在日粮中添加姜黄素后,NC、IC组仔猪空肠T-AOC、CAT的活力显著升高,NC组GSH含量显著增多,NC、IC组H2O2含量下降;IC组仔猪回肠CAT活力显著升高,NC、IC组仔猪回肠H2O2含量显著降低,且2 组T-AOC、T-SOD和GSH-Px的水平均呈现升高趋势。姜黄素可以改善氧化应激造成的抗氧化酶活力的降低。Wang Na等[27]的研究结果显示,姜黄素显著缓解H2O2诱导产生氧化应激而导致的SOD活力降低。Sivalingam等[17]研究表明,姜黄素显著升高肠道细胞CAT活力,对GSH-Px活力也有升高趋势,与本实验结果相似。由此可见,姜黄素对提高IUGR断奶仔猪肠道抗氧化功能具有一定的帮助作用。
有研究表明,姜黄素抗氧化机制是通过Nrf2-KEAP-抗氧化反应元件(antioxidant responsive element,ARE)通路来发挥其作用的[5,17]。Nrf2是外源性有毒物质和氧化应激的感受器,在参与细胞抗氧化应激和外源性有毒物质诱导的主要防御机制中发挥重要的作用[31]。KEAP是通过保护性蛋白DNA上游调节区的ARE来调控的[32-33]。当细胞受到刺激(包括氧化剂、抗氧化剂和化学预防剂)时会激活Nrf2因子,导致Nrf2从KEAP1上解离并转移到核上,从而增强抗氧化系统和II相解毒酶系统的相关基因表达,减少或消灭细胞受到的刺激[34]。主要的II相解毒酶系统包括GST、NQO1、HO-1、SOD、GSH-Px等[31,34]。本研究观察到IUGR断奶仔猪空肠Nrf2、KEAP1、SOD、GSH-Px1、GST、NQO1、HO-1、GCLM和TXNRD1基因的表达量显著降低;回肠CAT、SOD和GSH-Px1基因的表达量显著降低,NQO1和TXNRD1基因的表达量有所降低。近年来研究发现,与正常组相比,IUGR仔猪空肠黏膜中GSH-Px1、CAT和TXNRD1的mRNA表达量显著降低,HO-1表达量有所下降[35]。TXNRD1是唯一已知的将硫氧还蛋白保持在还原状态的酶,也是肠上皮细胞中的主要细胞内氧化还原系统,起到清除ROS物质的作用[36-37]。因此,抗氧化基因表达显著降低也可能反映IUGR仔猪肠黏膜的抗氧化能力受到干扰。本实验在日粮中添加姜黄素后,NC组空肠CAT、GST和NQO1基因的表达显著升高,回肠GST和TXNRD1基因的表达显著升高;IC组空肠Nrf2、CAT、SOD、GST和NQO1基因的表达显著升高,回肠Nrf2、GST、NQO1和TXNRD1基因的表达显著升高。Nrf2及其下游NQO1、SOD1和HO-1表达上调是Nrf2-ARE通路被激活的标志[38]。上述结果提示姜黄素可能通过激活Nrf2-ARE通路来缓解IUGR造成的仔猪肠道氧化应激和提高肠道抗氧化能力。
综上,IUGR会引起断奶仔猪肠道发生氧化应激,破坏抗氧化防御系统,而日粮中添加400 mg/kg姜黄素这一措施对缓解IUGR诱导的仔猪肠道氧化应激和提高肠道抗氧化功能有一定作用,也为改善动物和人类IUGR后代肠道氧化损伤提供新的依据。
猜你喜欢
首都食品与医药(2022年23期)2023-01-25
今日农业(2022年2期)2022-11-16
世界科学技术-中医药现代化(2021年9期)2021-12-31
河南畜牧兽医(2020年11期)2020-01-11
Medical Data Mining(2019年2期)2019-07-16
猪业科学(2018年5期)2018-07-17
中成药(2018年2期)2018-05-09
中成药(2018年3期)2018-05-07
中国塑料(2015年3期)2015-11-27
中国当代医药(2015年33期)2015-03-01
