
养殖与野外捕获大黄鱼躯干侧线系统的初步比较
2020-01-09 01:59林听听
海洋渔业 2019年6期
刘 鑫,张 东,林听听
(中国水产科学研究院东海水产研究所,农业农村部东海渔业资源开发利用重点实验室,上海 200090)
侧线是水生无羊膜动物用于感知周围刺激的感觉器官,存在于鱼类和两栖动物水生幼体中,主要功能是感知周围水流的低频(<200 Hz)振动,在鱼类趋流、洄游行为中发挥重要作用,同时还能协助视觉对远处物体定位,对鱼类的食物搜寻、避敌、产卵和集群等行为均有重要作用[1-3]。
侧线系统机械感受的基本单元是神经丘。根据神经丘在体表的位置,可将其分为表面和管道神经丘两大类[7]。管道神经丘常包埋于膜骨内或软骨质凹槽中形成头部侧线,在躯干部则位于侧线鳞或包裹在皮下,头部和躯干侧线均通过管道孔与外界水体接触。表面神经丘分布在鱼体表面,有时位于皮肤的突起上或凹槽内[4-7]。鱼类的侧线形态在具备种属特异性的同时,还随生活环境及生活习性的变化而改变,具环境适应性[8-9]。例 如,栖 息 在 溪 流 中 的 三 刺 鱼(Gasterosteus aculeatus)比水体底层或平静水域的同种个体具有更多的管道神经丘[10],栖息在缓流环境的鱼类常常具有发达的表面神经丘和不连贯的、甚至无侧线管道[11],如暗纹东方鲀(Takifugu obscurus)等[12]。
大黄鱼(Larimichthys crocea),隶属于硬骨鱼纲,鲈形目(Perciformes),石首鱼科(Sciaenidae),黄鱼属,因其肉质细嫩鲜美、营养丰富而颇受消费者的喜爱,曾是中国东海、黄海及南海的主要经济鱼类之一。20世纪70年代以来,由于过度捕捞,野生大黄鱼种质资源几近枯竭。自1985年大黄鱼人工育苗和养殖试验成功后,大黄鱼的人工养殖业迅速发展,不仅满足了市场需要,也为大黄鱼的增殖放流提供了技术条件[13]。为保护大黄鱼的种质资源,维护生态平衡,自1998年起,大黄鱼的人工放流在沿海多地展开[14]。
由于人工养殖环境和野外环境的水流、捕食与被捕食等生存压力差异很大[15-16],养殖大黄鱼和生活在野外环境中的大黄鱼侧线系统可能有差异,如果养殖大黄鱼的侧线系统并不适合野外生存,那么养殖大黄鱼被放流到野外环境后,其食物搜寻、避敌、产卵和集群等行为可能受到影响,从而危及生存。因此,研究人工养殖和野外捕获大黄鱼侧线系统的差异不仅有助于研究大黄鱼侧线系统发育生态学,还可为进一步研究大黄鱼行为、评估人工放流效果奠定基础。本文对人工养殖和野外捕获大黄鱼的侧线器官进行了形态学的初步研究,探索大黄鱼侧线系统适应性进化的方向,以期为大黄鱼增殖放流群体的选育提供理论依据。
1 材料与方法
1.1 实验用鱼
实验用养殖大黄鱼幼鱼[体长(14.1±1.3)cm、体质量(44.0±7.2)g]和仔鱼于 2016年购自福建省宁德市某水产有限公司。野外捕获的大黄鱼体长(14.2±2.7)cm、体质量(43.4±8.0)g,于2016年采自吕四渔场。
1.2 实验方法
养殖大黄鱼自孵化后每24 h取材1次,直至30日龄。每次取6尾样品,经2.5%戊二醛溶液固定24 h后,移入梯度乙醇溶液中脱水、经临界点干燥,JSM6380-LV扫描电镜观察并拍照,用于对侧线器官的观察与分析。
养殖大黄鱼和野外捕获的大黄鱼各10尾,经0.003%的3-氨基苯甲酸乙酯甲基磺酸盐(MS222)溶液麻醉后,以10%的中性甲醛溶液固定1~7 d。其中5尾浸泡在0.5%KOH溶液中,直至样本体表色素消除,然后以0.1%的亚甲基蓝溶液对样品的躯干侧线管进行注射,观察侧线的排布并拍照。另取5尾鱼的躯干侧线皮肤,切割为0.3 cm×0.3 cm的组织块,用于扫描电镜的观察,实验方法同前述。
用于组织学观察的样品经梯度乙醇溶液脱水、二甲苯透明、石蜡包埋后制备厚度为5~6 μm的连续切片,经H.E.染色,中性树胶封片,镜检后拍照。
1.3 统计方法
采用T检验比较养殖和野外捕获大黄鱼神经丘数目的差异,数据分析采用统计软件SPSS 19.0进行。
2 结果与讨论
2.1 大黄鱼侧线器官的形态观察
大黄鱼的侧线系统由管道神经丘和表面神经丘两类感受器组成(图1-a,b)。神经丘的感觉斑上分布有毛细胞(图1-c,d)。毛细胞常形成的纤毛簇,由一根较长的动纤毛和一簇高度从高到低排列的静纤毛组成(图1-e,f)。大黄鱼的管道神经丘位于侧线管道腔内的皮肤表面,2个侧线管道开口之间有1个神经丘,管道神经丘的感觉斑为椭圆形,其长轴与鱼体的体轴垂直,其上着生感觉毛,毛细胞的顶端常具粘液团(图1-a,c)。大黄鱼的表面神经丘多具有圆形或近圆形的底座,底座高于表皮层,神经丘的顶端具成簇的毛细胞(图1-b,d,f;图2)。
0~8日龄仔鱼的神经丘全部为表面神经丘,分布在头部、躯干和尾鳍,头部的神经丘分布在眶上、眶下、颞上、耳后、前鳃盖以及下颌,表面神经丘的数目为3~12个。30日龄幼鱼鳃盖处的表面神经丘位于凹陷的皮肤表面(图3-a),此时躯干部侧线管道闭合,具备管道神经丘(图2-a),头部表面神经丘的数目为(24.6±2.9)个。在70日龄幼鱼的头部,除仅在眶前近吻端的位置观察到表面神经丘以外,其他各处均未观察到表面神经丘,躯干侧线管开口于侧线鳞上,侧线鳞片上具有表面神经丘(图3-b)。
2.2 养殖大黄鱼与野外捕获大黄鱼躯干侧线系统的比较
大黄鱼身体左右两侧各具有1条躯干侧线管,养殖与野外捕获大黄鱼的躯干侧线均自鳃盖后缘沿脊椎骨的方向,延伸至尾柄的中点(图4-b,c)。侧线管无分枝,通过侧线鳞上的管道开口与外界接触(图4-a,c)。躯干侧线上的管道神经丘位于由侧线鳞组成的管腔内,每个鳞片处着生1个神经丘(图5-a,b),养殖大黄鱼的管道神经丘与野外捕获大黄鱼的管道神经丘结构类似,均由毛细胞和支持细胞组成(图5-c,d)。养殖大黄鱼躯干单侧侧线的管道神经丘数目为(54.7±1.7)个,野外捕获大黄鱼单侧躯干部有(55.0±3.3)个管道神经丘,二者无统计学差异(t=0.128,P=0.45)。大黄鱼的侧线鳞边缘的开口处有数个表面神经丘,就单个鳞片上表面神经丘的数目而言,养殖大黄鱼有(7.60±2.57)个神经丘,野外捕获大黄鱼有(4.90±1.76)个神经丘,二者之间存在显著性差异(t=2.11,P=0.02)。
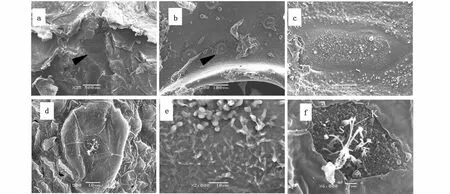
图1 扫描电子显微镜下大黄鱼的神经丘Fig.1 Scanning electron micrographs of neuromasts in Larimichthys crocea
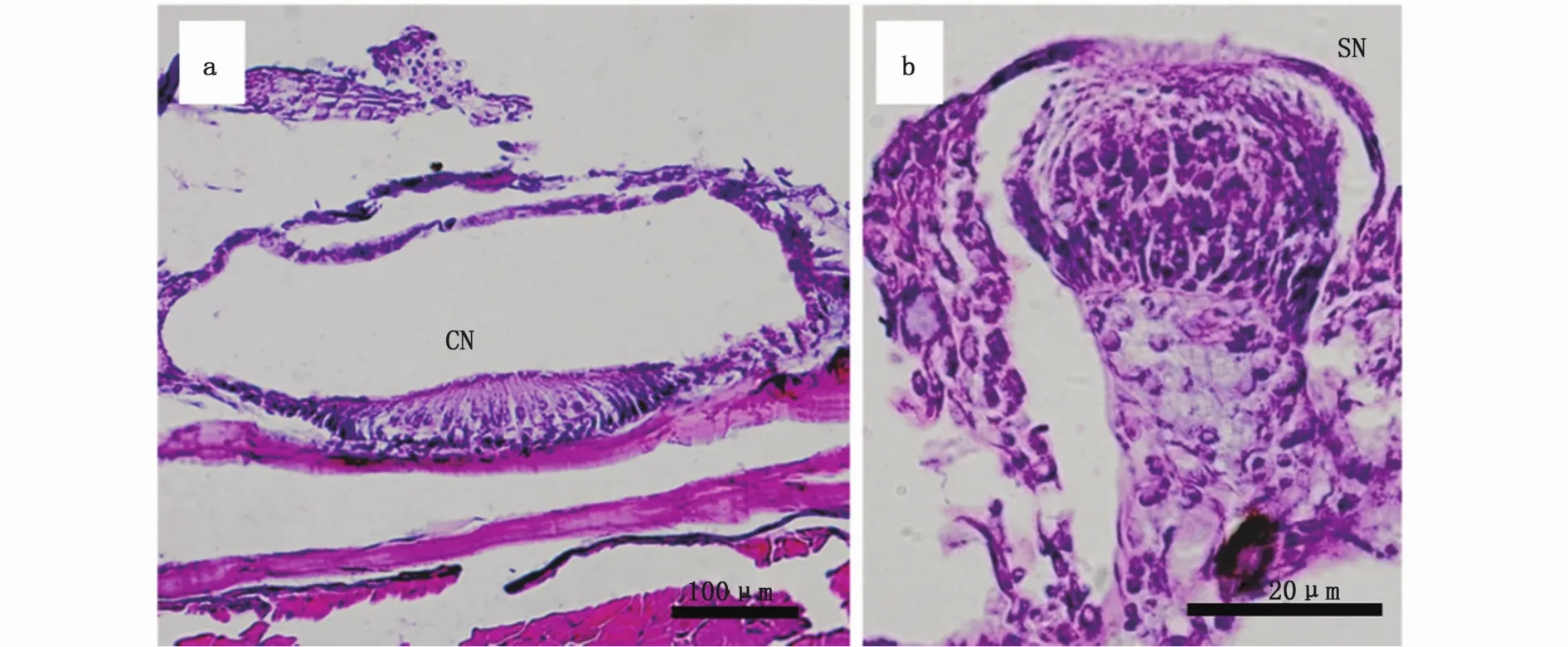
图2 神经丘的结构Fig.2 Neuromast structure of Larimichthys crocea
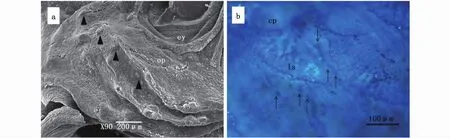
图3 大黄鱼的表面神经丘Fig.3 Superficial neuromast of Larimichthys crocea
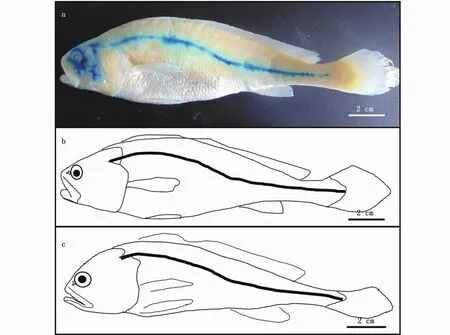
图4 大黄鱼的躯干侧线Fig.4 Trunk lateral line of Larimichthys crocea

图5 养殖大黄鱼与野外捕获大黄鱼的躯干侧线管道及管道神经丘Fig.5 Lateral line canal and neuromasts of cultured and wild-caught Larimichthys crocea
2.3 讨论
大黄鱼的侧线感受器分为表面神经丘和管道神经丘两种,二者均具有感觉斑,感觉斑的表面分布有毛细胞。头部神经丘主要分布在眶上、眶下、颞上、耳后、前鳃盖以及下颌,符合辐鳍亚纲鱼类头部侧线系统的基本分布模式[6]。与大部分硬骨鱼[8,15-16]类似,大黄鱼仔鱼的神经丘全部为表面神经丘,从鳃盖处表面神经丘的位置变化推测,大黄鱼头部侧线管道上皮凹陷,先形成半开放的管道,然后逐渐闭合形成侧线管。
研究表明,表面神经丘与管道神经丘的相对丰度除具有种属特异性外,也是鱼类对栖息地水流环境适应的结果。栖息于底层缓流或相对平静水域的鱼类常具有不完整的侧线管,体表仅具较高比例的表面神经丘[11]。栖息于静水中的鲈鱼(Percidae)具较高比例的表面神经丘、较宽的侧线管道;而喜好急流生境的种类常常具有狭窄的侧线管道[18-21]。养殖大黄鱼与野外捕获大黄鱼的躯干部侧线分布、侧线管道形式均类似,说明其管道侧线的位置保守,虽然生活在不同的人工与自然环境中,二者躯干部侧线管道的分布模式并无明显的差别,推测养殖大黄鱼与野外捕获大黄鱼的栖息地水流环境可能类似。
在人工养殖条件下,大黄鱼多生活在缓流的近岸网箱或室内水泥池中[12,22],存在高密度放养的情况,由于表面神经丘主要感知鱼体周围近场的机械振动信号[4-7],养殖大黄鱼需具备足够多的表面神经丘,对近距离的物体引发的振动信息进行整合,以适应高密度的养殖水体。BROWN等[23]对于养殖虹鳟(Oncorhynchus mykiss)与野生种类侧线形态做比较研究发现,养殖个体表面神经丘的密度明显低于野生个体,而养殖虹鳟的野外生存能力较弱,因此,低密度的表面神经丘被认为是侧线系统异常或退化的表现,侧线系统的退化会减弱与之相关的游泳、觅食、避敌等行为能力,进而导致养殖虹鳟在野外的存活率低。野外捕获大黄鱼躯干部单片鳞上的表面神经丘数目少于养殖大黄鱼,推测可能是由于自然生境与人工养殖环境的选择压力不同,导致野外的大黄鱼侧线系统出现异常或退化,引起机械感受功能的减弱甚至缺陷,影响种群的生存。
当茴鱼(Thymallus thymallus)被投放到自然环境中后,它们常停留在放流地附近的水底、低流速水域[24-25]。人工养殖的虹鳟在洄游起始阶段的死亡率为30% ~40%,而野生个体的死亡率仅为7% ~13%;有26% ~40%的养殖个体进入大洋,明显低于野生个体(71%~84%)[26]。人工养殖的鱼类在放流到野外环境后,在游泳、避敌、感知能力等方面均弱于野生个体。由此推测,养殖大黄鱼被直接放流到自然生境中后,也可能由于侧线的机械感受功能与野生鱼不同,而表现出与野生鱼生存能力不同的情况。
现有的研究结果显示,与野生的亲本相比,官井洋的养殖大黄鱼呈现出眼大、吻短、体高的形态特征,而这些形态结构的变化都极有可能引起相应感觉器官功能的变化[27]。本实验中野外捕获大黄鱼可能来自于大黄鱼的野生群体,也可能来自从网箱中逃逸的养殖鱼群体,无论源自何种群体,本结果均表明,在不同的生存环境下,大黄鱼的躯干侧线系统会发生变化,该变化主要体现在侧线鳞片的表面神经丘数目上。今后可针对大黄鱼侧线系统的表面与管道神经丘的相对比例,全面阐释两种生境下侧线结构的异同。此外,不同生境下大黄鱼侧线系统的功能是否存在差异,如趋流性、食物搜寻、躲避敌害等能力是否不同,仍需进一步的实验验证。今后可针对大黄鱼躯干侧线系统适应性变化所需的条件、时长以及变化过程开展研究,明确其适应性进化的方向,为大黄鱼增殖放流群体的选育提供理论依据。
猜你喜欢
海洋开发与管理(2022年10期)2022-11-11
军事文摘(2022年8期)2022-11-03
体育科技文献通报(2022年4期)2022-10-21
辽河(2022年5期)2022-07-07
体育科技文献通报(2022年5期)2022-06-05
农产品市场周刊(2021年17期)2021-09-28
小学科学(学生版)(2021年3期)2021-04-13
哈哈画报(2021年11期)2021-02-28
诗选刊(2019年9期)2019-11-20
当代水产(2019年6期)2019-07-25
