
Effect of Probiotics on Juvenile Tachypleus tridentatus Gut Microbiota
2022-06-14 06:30XIONGKaiMIAOFengzeWAIHOKhorLIUXimeiFANGJamesHEJinfengHUMenghongandWANGYouji
XIONG Kai, MIAO Fengze, WAIHO Khor, LIU Ximei, FANG James K. H.,HE Jinfeng, HU Menghong, *, and WANG Youji
Effect of Probiotics on JuvenileGut Microbiota
XIONG Kai1), 2), MIAO Fengze1), 2), WAIHO Khor3), LIU Ximei1), 2), FANG James K. H.4),HE Jinfeng5), HU Menghong1), 2), *, and WANG Youji1), 2)
1),,,201306,2),,201306,3),,,4),,,999077,5),536000,
The population of tri-spine horseshoe crabs (HSCs)has sharply decreased in recent decades. Do- mestication and manual releasing are now used to recover it. In order to accelerate the growth ofin indoor environment, in this study, gut microbiota of 2nd instarwas supposed to be improved by putting probiotics in water environment and thus promoted the health and growth of the host. Using 16S rRNA sequencing of gut samples, we examined the intestinal micro- biome of juvenile HSCs and studied the effect of probiotics. Results showed that there was no significant difference between groups fed with high and low concentrations (1×107, 1×1012CFUmL−1) ofand high concentration (1×1012CFUmL−1) of, and no significant difference compared with the control group, but the diversity ofgroup with low con- centration (1×107CFUmL−1) was significantly higher than those of other groups. Microbial-mediated functions predicted by PICRUSt showed that ‘Cellular Processes’, ‘Metabolism’ and ‘Organismal Systems’ pathways significantly increased in groups ofandgroup with low concentration. The results showed that two types of probiotics with different concentrations have other effects on the structure, richness and function of intestinal microbes. By searching the literature and conducting pre-ex- periments, we forecasted the short time of breeding under the probiotic environment may not have much effect. The present study for the first time investigated the impact of probiotics on gut microbiota of juvenile HSCs and found them may benefit for the growth and health of HSCs.
; probiotic; gut microbiota; diversity; richness
1 Introduction
Horseshoe crabs areone of the most ancient marine organisms in the nature. They first appeared in the Devo- nian period of Paleozoic, and are known as ‘living fossils’ because their morphological structures have not changed much since then (Sekiguchi and Nakamura, 1979; Shuster, 1979; Chen, 2015). Tachypleus amebocyte lysate (TAL), which is made using the hemolymph of,is internationally recognized as the most sen- sitive bacterial endotoxin detection product (Hong.,2016). At present, there are only three genera and four spe- cies of horseshoe crabs in the world.is main- ly distributed in Guangdong, Guangxi and Fujian provinces and region in China (Carmichael and Brush, 2012). Un- fortunately, habitat destruction, production of TAL, and overfishing have caused a sharp decline in thepopulation (Shin, 2007; Liao, 2019), and it is now classified as a Class II national protected species in China.
‘Probiotics’ are active microorganisms that contribute to the balance of gut microbes (Dai and Chen, 1995). Probio- tics can significantly induce intestinal flora to produce a variety of metabolites-volatile short-chain fatty acids, which play key roles in maintaining intestinal health of aquatic species. Competition inhibition is an important mechanism of probiotics, and also the main viewpoint of microecolo- gical theory. Through competition for fixed sites, probiotics reduce the number and density of pathogenic bacteria, thereby contribute to disease prevention (Tian, 2011; Barton and Mary, 2014). In aquaculture, feeding probio- tics such asand its mixture can improve the activities of superoxide dismutase(SOD) and catalase (CAT), as well as total antioxidant capacity (T-AOC) inblood. It also can reduce malondialdehyde(MDA) content,improve lysozyme activity and antioxidant capacity, and increase immune function of the organism (Wang,2021).can regulate the normal intestinal flo- ra of an organism, maintain microecological balance, en- hance its immune function, improve disease resistance by producing antibacterial metabolites and inhibit the growth of. Therefore,andwere selected as experimental strains in this experiment.
The decline in the population ofis expect-ed to exert ecological and social impacts. Raising offspringto increase the population is considered as an option for increasing the population of,which will fur- ther support sustainable resource management and research activities. Hatchery and culture techniques for the species have been developed since the late 1980s, and large-scale farming is common in Japan (Iwaoka and Okayama, 2007). Previous studies mainly focused on the type of feed and feeding conditions to maximize the growth and survival ofjuveniles (Kwan., 2017). Little is knownabout the practicality and effectiveness of probiotics on the gut microbiota and the health of horseshoe crabs. Miao. (2020) compared the intestinal microbes of the first and second instars of, indicating that ecdysis ex- erted changes on the intestinal microbes of,andandwere genus-level biomarkers after molting.is a kind of pathogenic bacteria and probiotics can reduce it. In the early stage, antibiotics were used as a reagent to eliminate pathogenic bacteria and achi- eved better results, which was later found that long-term use of antibiotics would destroy the intestinal ecological balance and lead to drug resistance of bacteria (Hashemi and Davoodi, 2011; Looft, 2014). Since the prospe- rity of biotechnology, probiotics are more beneficial for the development of aquaculture, especially for feeding ap-plications and artificial culture technology (Yao, 2020).Therefore, the purpose of this study was to investigatewhether probiotics (and) hada positive effect on intestinal microbes from the second in- star to the third instar in a 21-day culture experiment. The results of this study provide a theoretical basis for the ef- fect of probiotics on crustacean intestinal microbiome du- ring instar developmental stages and spearhead further de-velopment of the artificial breeding technology of juvenile.
2 Materials and Methods
2.1 Experimental Conditions and Animals
Juveniles ofwere artificially raised in the laboratory (Beihai Xinglong Biological Products Co., Ltd.). Wildwere stimulated to lay eggs and ferti- lized by simulating tidal changes during the breeding pe- riod. Egg incubation period was approximately 45–50d.Newly hatched first instars were raised in a glass tank (1.5m×0.7m×0.3m) with circulating water system at Shang- hai Ocean University (Shanghai, China). Six baskets (0.35m×0.2m×0.1m) covered with fine sands were placed in the glass tank, and each basket contained 100 first instars. This experiment was conducted in June, and the experimen- tal conditions were maintained at temperature 29–30℃, pH 8.1–8.2, salinity 19–32, dissolved oxygen 6.0–8.0mgL−1, and photoperiod 12h:12h (darkness:light). Approxi- mately one third of the total water volume was changed every three days. No feeding was required during the first instar stage. After about 60d, the 1st instarmolted to become the 2nd instarThen they were transferred into experimental tanks (0.25m×0.165m×0.16m, 6.6L), with 10 instars per tank. Five treatments were used in this study, including control (Ctr) without any treatments, low concentration of(1×107CFUmL−1) (Lbs), high concentration of(1×1012CFUmL−1) (Hbs), low concentration of(1×107CFUmL−1) (L1), and high concentration of(1×1012CFUmL−1) (H1). One tank represent- ed one treatment replicate, and triplicates were conducted for each treatment. We once took out the intestines ofand observedthrough microscope. So(approximately 1g) was fed to juvenile horseshoe crab once a day. We mixedand probiotics to feed horseshoe crabs, and changed 1000mL of configured sea- water after 2h of feeding. The culture experiments were conducted for a period of 21d.
2.2 Sample Collection
Juvenileof second instar were collected, dried on a test paper, disinfected with 70% alcohol, and re-washed with distilled water. After drying on a test pa- per, the complete intestines of 10 instars were collected in- dividually, snap frozen with liquid nitrogen and stored at −80℃ until further analysis (Fig.1).
Fig.1 The gut of 2nd instar T. tridentatus. Scale bar length is 0.25cm.
2.3 DNA Extraction and Sequencing
The intestinal tissue specimens were crushed, and 0.4g of the sample powder was placed in a 2mL microcentri- fuge tube. Subsequently, DNA was isolated with a Mag- Pure Soil DNA LQ Kit (Magen, Guangdong, China) fol- lowing the manufacturer’s instructions. According to the method of Kumar. (2011), the target area of V3-V4 of bacterial 16S rRNA was used to compare the gut mi- crobiome of juvenile HSC of different groups. Sequenc- ing was performed on an Illumina NovaSeq6000 (Illumi- na, San Diego, CA, USA) at Shanghai OE Biotech. Co., Ltd. (Shanghai, China).
2.4 Data and Bioinformatic Analysis
Data are presented as mean±standard error (SE) for each group. Trimmomatic software was used to preprocess the paired-end reads (Bolger, 2014) for the detection and cutting off of ambiguous bases (N). Sequence denoising was further performed to remove reads with unclear se- quence, homology or less than 200bp. QIIME software (ver- sion 1.8.0) was used to retain 75% or more of the Q20 base reads (Caporaso, 2010). Cleaning reads were sequen- tially removed and clustered, and operational taxonomic units (OTUs) were generated using VSEARCH software with similarity truncation of 97% (Rognes, 2016). QIIME package was used to select a representative read- ing for each OTU. Unweighted Unifrac principal coordi-nate analysis (PCoA) and phylogenetic tree construction were performed using Unifrac distance matrix generated by QIIME software. Sequencing and analysis of 16S rRNA amplicon were performed by OE Biotechnology Co., Ltd. (Shanghai, China).
3 Results
3.1 Operational Taxonomic Units (OTUs) and Diversity Analyses
After 16S rRNA sequence analysis, Ctr (control), Hbs (high concentration of), Lbs (low concen- tration of), Hl (high concentration of) and Ll (low concentration of) wereanalyzed. The Alpha diversity analysis showed that Ctr, Hbs, Lbs and Ll dropped abruptly on the OTU rank graph while Hl smoothly declined, which indicatedthat the bio- diversity of Ctr, Hbs, Lbs and Ll were not significantly different while the biodiversity of Hl was significantly high- er than the other four groups (Table 1, Fig.2).

Table 1 Statistics of the Alpha index of intestinal microbiome and water microbiome
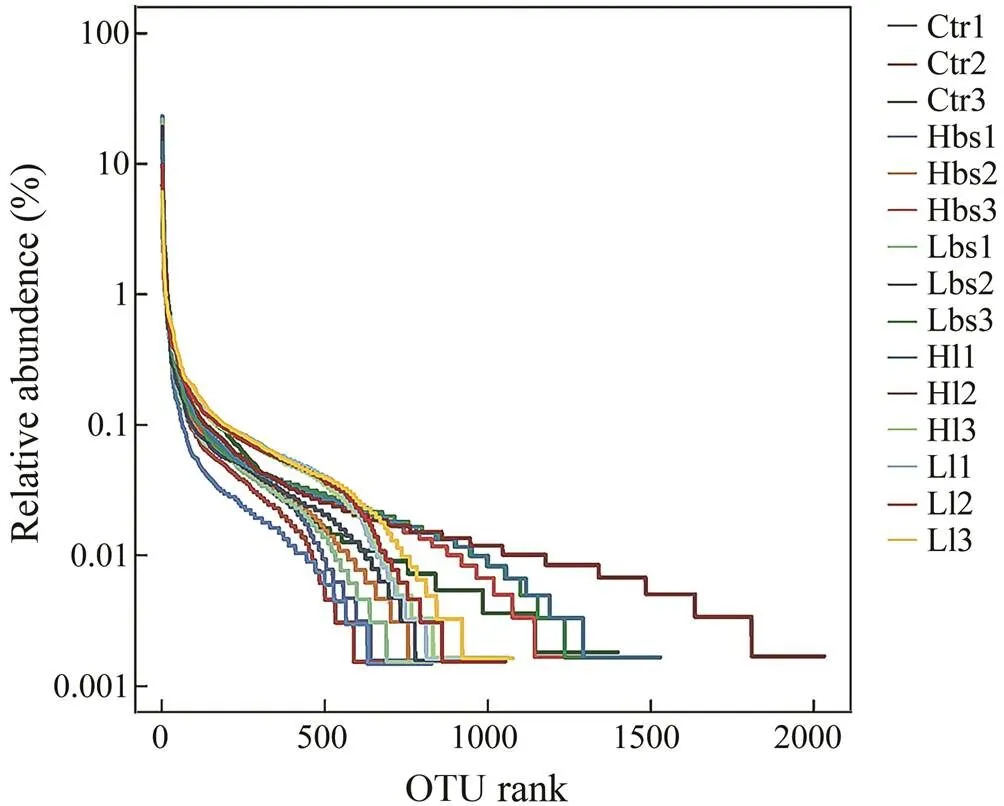
Fig.2 Statistics of OTU rank and relative abundance of Alpha analysis. Ctr data were collected before the 21-d experiment.
According to Alpha analysis, the chao1 (Fig.3a) indices of Ctr, Hbs, Lbs, Hl and Ll had no significant difference, but the species richness of Hl was significantly higher thanthose of other groups. Shannon (Fig.3b) and Simpson (Fig.3c) indices of Ctr, Hbs, Lbs, Hl and Ll were not significantly different, but the biodiversity of Hl and Ll were significant- ly higher than those of other groups.
PCoA analysis of composition of three samples (Ctr, Hbs and Lbs) were mostly overlapped, and the difference be- tween Hl and Ll samples was also not obvious. However, the bacterial compositions of Ctr, Hbs and Lbs were sig- nificantly different with Hl and Ll samples (Fig.4).
3.2 Composition Analysis at Phylum Level
Cluster analysis was performed on the basis of OTUs and sequencing data to obtain a histogram of the micro- bial community of Ctr, Hbs, Lbs, Hl and Ll at the phylum level, showing the columnar distribution of the relative abun- dance of the top 15 phyla (Fig.5). In general, the intestinal microbiota of the second instars was dominated by micro- bes from the category of,,,,,,,,,,,,,and. In the five groups of the HSC gut microbiome (Ctr, Hbs, Lbs, Hl and Ll),,andwere always the dominant phyla. As for Ctr, Hbs and Lbs,was the most dominant phylum (56.6, 48.0% and 56.5%), fol- lowed by(18.5%, 26.7% and 22.6%) and(18.4%, 19.6% and 14.6%). As for Hl,(31.5%),(31.1%) and(28.0%) were still the three most advantageous cate- gories, but the proportion ofdecreased and the proportion ofincreased compared with other groups. As for Ll,was replacedas the most dominant phylum (55.3%) followed by(13.0%),(12.2%) and(10.5%). Besides, it is worth noting that the propor- tion ofin Ll had increased. The Ctr and Lbs abundances of other microorganisms were <7%. The Hbs abundances of other microorganisms were <6%. The Hl and Ll abundances of other microorganisms were <10%.
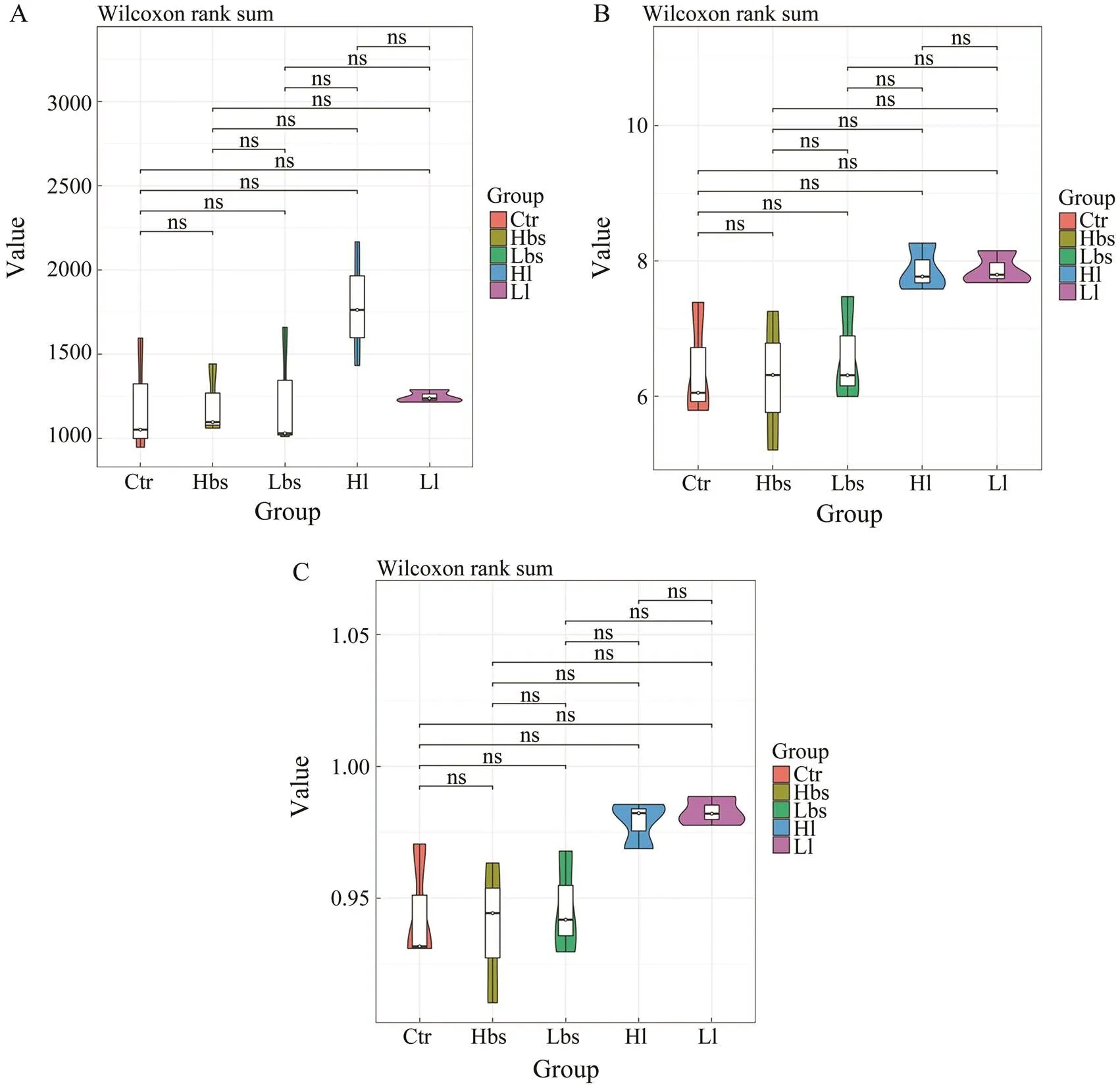
Fig.3 Alpha metrics of intestinal microbiome: Chao1 index, and Shannon diversity.

Fig.4 Principal component analysis of intestinal micro- biome in HSC.
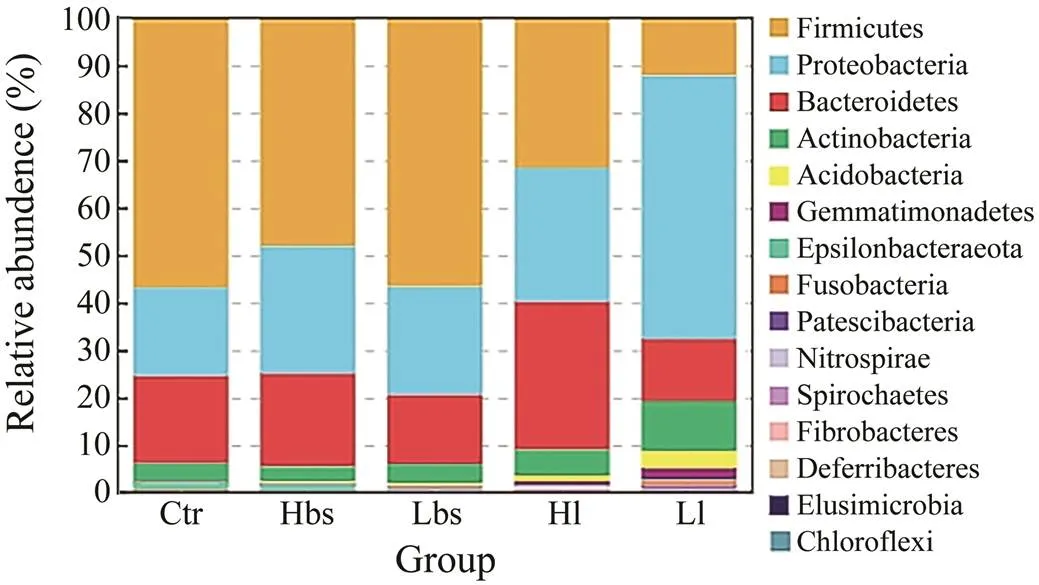
Fig.5 At the phylum level, the relative abundance of in- testinal bacteria in 5 groups were in the top 15.
3.3 Unique Biomarkers Detected in Juvenile HSC Intestines
The LEfSe analysis in Ctr, Hbs, Lbs and Ll revealed (Fig.6) thatwas the genus-level biomarker in Ctr.,,andwere the genus-level biomarkers in Hbs; Gemmatimo- nadetes and Nitrospirae were phylum-level biomarkers in Lbs; and,,,andwere genus-level bio- markers in Ll.
3.4 Function Prediction
The KEGG pathways related to the intestinal function of the second instars changed to a certain extent after add- ing different concentrations and different types of probio- tics. As for Ll and Lbs, ‘Cellular Processes’ pathways, ‘Me-tabolism’ pathways and ‘Organismal Systems’ pathways were found more significantly different than those in Ctr, Hbs and Hl. In addition, ‘Replication and Repair’ pathways,‘Immune System’ pathways and ‘Environmental Adapta- tion’ pathways were found more abundant in Lbs. ‘Envi- ronmental Adaptation’ pathways, ‘Transport and Catabo- lism’ pathways, ‘Cell Growth and Death’ pathways, ‘Energy Metabolism’ pathways and ‘Metabolism’ pathways were found more abundant in Ll (Fig.7).
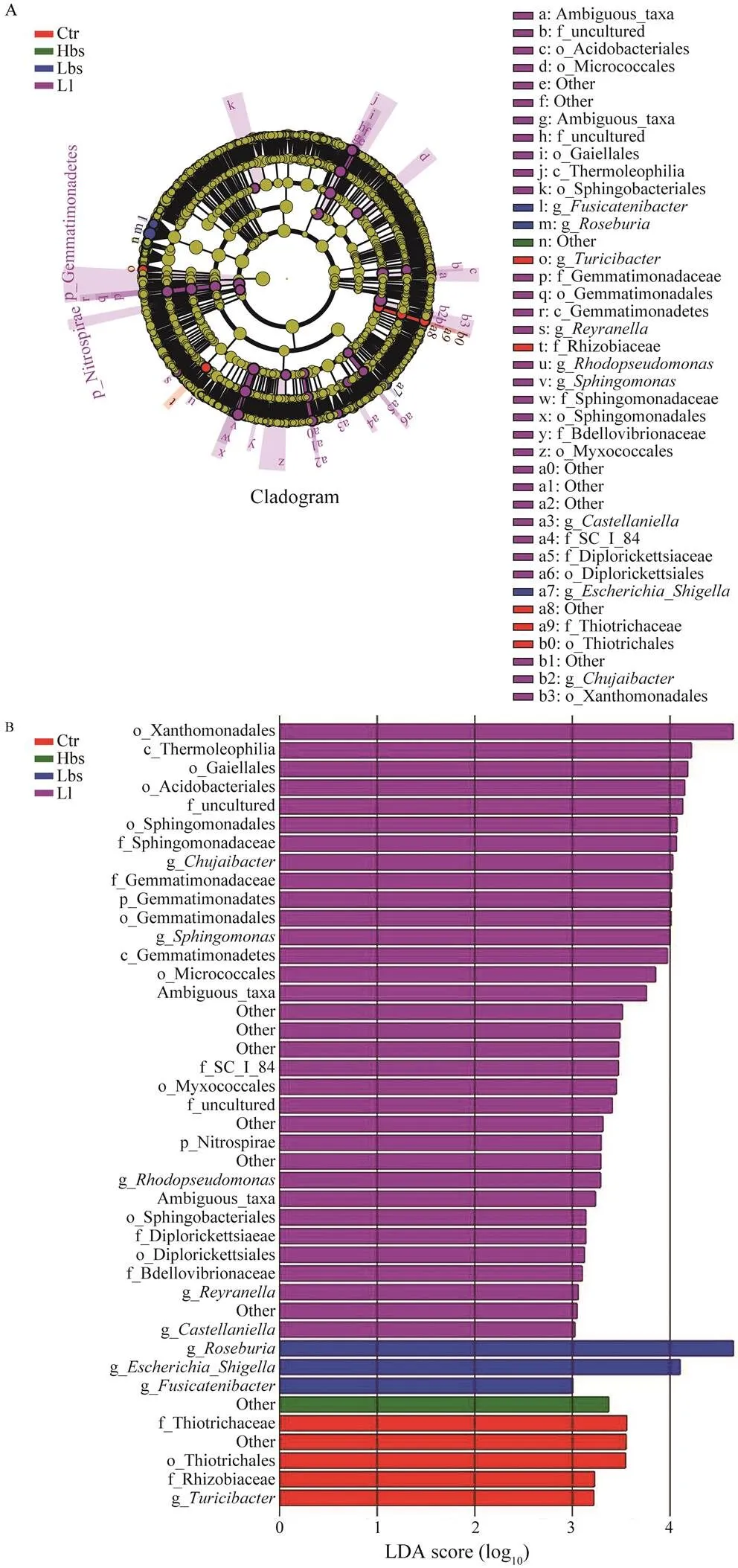
Fig.6 LEfSe histogram of intestinal flora of Ctr (control), Hbs (high concentration of Bacillus subtilis), Lbs (low concentration of Bacillus subtilis) and Ll (low concentration of Lactobacillus) showed the log-Transform LDA score of bacterial groups appraised by LEfSe analysis (log-Transform LDA score 3.0 was the threshold). The single characters before the underlines are abbreviations: p, phylum; c, class; o, order; f, family; g, genus.
4 Discussion
The composition of gut microbes is influenced by manyfactors, such as physiology, genetic inheritance, geography, life history and living environment (Yang and Shao, 2013).The formation and stability of the intestinal environment will not only affect the growth or death of the host, but alsoimprove or reduce metabolic transformation, immunity and many other important physiological functions (Sun, 2020).The research of intestinal microbes has been a very popular topic in other aquatic organisms, but the research on the intestinal microbes of HSCs needs more attentions.
In recent years, probiotics have been widely favored by researchers due to their safety and ability to inhibit patho- genic bacteria. In the present study, the intestinal micro- bes of the second instars of HSCs are more abundant after the introduction of,both inhigh and low con- centrations (1×107, 1×1012CFUmL−1). It is worth of not- ing thatshows significantly higher propor- tion than the other three groups (Kämpfer, 2010). However,the proportions ofin the groups treated with high and low concentrations ofwere similar to that in Ctr.is a biologically ac- tive substance that is more abundant than other microor- ganisms and has antibacterial activity as the main produ- cer of antibiotics (Kämpfer, 2010).The results indicate that the introduction ofto the living environment of second instars can increase the proportion of actinomy- cetes in the intestinal microbes and reduce the proportion of pathogenic bacteria. This slight optimization may im- prove the health of the hosts’ intestinal microbial commu- nity.
In the expression of the intestinal microbial richness, the intestinal microbes of the 5 groups of HSCs change with no significance after 21 days, but the intestinal microbial abundance of the two groups after treatment withwas still slightly higher than those of the other three groups. Firstly, the intestinal flora of aquatic animalswill be affected by many factors, not only genetic and phy- siological differences, but also environmental factors (Ni- cholson., 2012). Research reports indicate that the number of bacteria in the intestine of freshwater fish is basically 105–108cfug−1, while the number of bacteria in the intestine of marine fish is 106–108cfug−1. Because of the great amount of intestinal microbes, the change of in- testinal flora is a relatively long process (Sugita, 1983; Guo, 2019).
In the present research, the microbial composition of the intestinal tract has been greatly changed after feedingpro- biotics. KEGG prediction indicates that plenty of bacte- rial KEGG pathways significantly changed after feeding with low concentration of(Lbs) and low concentration of(Ll). For example, the ‘Cel- lular Processes’, ‘Metabolism’ and ‘Organismal Systems’ significantly increased in Lbs and Ll, but then significant- ly debased in Ctr, Hbs and Hl. Additionally, significant im- provement of these pathways indicates that the metabo- lism and digestion capacity of juvenile HSCs can be im- proved by changing the breeding environment and adding appropriate amounts of probiotics, which can promote the growth and development of HSCs.However, different con- centrations of the same type of probiotics may also lead to various results. The ‘Replication and Repair’ and ‘Immune System’ pathways have been significantly improvedin Lbs, which indicates that feeding an appropriate amount ofcan increase the immune system of ju- venile HSCs. On the other hand, ‘Digestive System’, ‘Trans- port and Catabolism’, ‘Energy Metabolism’ and ‘Metabo- lism’ pathways significantly increase in Ll, which indicates that feeding an appropriate amount ofcan increase metabolism and digestion of juvenile HSCs. Aqua- tic crustaceans will soon be affected by conditions such as temperature, light, and salinity when they molt. They will also be affected by conditions such as energy accumula- tion and adequate nutrition. When the accumulation of energy and nutrients is not sufficient, the time between molting periods will be prolonged, and the growth rate and weight gain rate of molting will decrease, even resulting death finally.Therefore, the present study shows that pro- biotics can improve digestion and metabolism which may accumulate enough energy for the next molting of horse- shoe crabs and increase survival rate, which will be help- ful for the follow-up studies to explore whether the molt- ing cycle can be relatively reduced to achieve faster growth.
According to the result in the present study,has a greater impact on the intestinal microbes, whilehas less impact on the intestinal microbes. According to the LEfSe analysis,andwere phylum-level biomarkers ingroups.is a kind of gram-nega- tive bacteria, which is related to the process of eliminat- ing pathogenic bacteria, such as inhibiting the growth of pathogenic bacteria (Zeng, 2021).is a ge- nus-level biomarker ingroups, and a con-ditional pathogen that can reduce nitrate, meaning once the intestinal environment undergoes a major change it may become a pathogen (Shinohara, 2019).
In the present study, we find that two types of probio- tics with different concentrations showed other effects on the structure, richness and function of intestinal microbes. Li(2005) studied on the effect ofprepara- tions on the intestinal microbes of, showingthat the longer the cultivation time, the more en- ergy the microbes use. Wang(2009) usedto conduct experiments on the three common dominant microalgae in prawn farming ponds, showing that the in- hibitory effect ofon the three microalgae was obvious, but microalgae would slowly appear after stop- ping the addition ofBoth experiments changed the concentration of probiotics and both showed that a higher concentration cannot have a better improve- ment. Similarly, our results show that the high concentra- tion ofis not as effective as the low concen- tration on the second instars. In addition, the result withtreatment is similar to the control group. Asneeds long period to have a function, our breeding cycle is too short to have a significant result. The low concentration ofin the present study had a positive effect on the intestinal microbial richness, community distribution and predicted function of juvenile HSCs. We speculate that selecting the appropriate types of probiotics and the appropriate dosage for delivery may be an effective way to improve the health and growth of juvenile HSCs. Therefore, on the basis of our research, fur-ther research can be carried out, such as selecting more strains or setting up different breeding cycles to find the optimal solution for probiotics to improve the health and growth of juvenile HSCs.
Acknowledgements
This study was supported by the National Natural Sci- ence Foundation of China (No. 31872587), the CAS Key Laboratory of Coastal Environmental Processes and Eco- logical Remediation, and the YICCAS Grant (No. 2020 KFJJ11).
Barton, M. D., 2014. Impact of antibiotic use in the swine in- dustry.,19: 9-15, DOI: 10. 1016/j.mib.2014.05.017.
Bolger, A. M., Lohse, M., and Usadel, B., 2014. Trimmomatic: A flexible trimmer for Illumina sequence data., 30: 2114-2120, DOI: 10.1093/bioinformatics/btu170.
Caporaso, J.G., Kuczynski, J., and Stombaugh, J., 2010. QIIME allows analysis of high-throughput community sequencing data.,7: 335-336, DOI: 10.1038/nmeth.f.303.
Carmichael, R.H., and Brush, E., 2012. Three decades of horse- shoe crab rearing: A review of conditions for captive growth and survival.,4: 32-43, DOI: 10.1111/ j.1753-5131.2012.01059.x.
Chen, Z.B., Fan, H.Q., Liao, Y.Y., Qiu, G.L., Xie, H.L., and Lin, W.Y., 2015. A living animal facing a dilemma-horseshoe crab.,67: 3, DOI: 10.3969/j.issn.0368-6396.2015.03.015 (in Chinese with English abstract).
Dai, Z.J., and Chen, Z.P., 1995. Probiotics use prospects., 1995 (1): 46-47 (in Chinese).
Guo, Q.Q., Hu, J.N., Wei, J.S., and Zhang, H., 2019. Study me- thod of intestinal microflora on aquatic animal., 125 (1): 17-20 (in Chinese with English abstract).
Hashemi, S. R., and Davoodi, H., 2011. Herbal plants and their derivatives as growth and health promoters in animal nutri- tion., 35: 169-180, DOI: 10.1007/s11259-010-9458-2.
Hong, J., Hu, J. Y., and Ke, F., 2016. Experimental induction of bacterial resistance to the antimicrobial peptide Tachyplesin I and investigation of the resistance mechanisms., 60: 6067-6075, DOI: 10.1128/AAC. 00640-16.
Iwaoka, C., and Okayama, T., 2007. Public awareness and com- munity-based conservation for the horseshoe crab at Saikai Na- tional Park in Nagasaki Prefecture, Japan. International Sym- posium on the Science and Conservation of Horseshoe Crabs. Dowling Coll, Oakdale, NY, 571-575.
Kämpfer, P., 2010. Actinobacteria. In:. Timmis, K. N., ed., Springer, Berlin, Heidelberg, DOI: 10.1007/978-3-540-77587-4_133.
Kumar, P. S., Brooker, M. R., Dowd, S. E., and Camerlengo, T., 2011. Target region selection is a critical determinant of com- munity fingerprints generated by 16S pyrosequencing., 6 (6): 1-8, DOI: 10.1371/journal.pone.0020956.
Kwan, B. K. Y., Chan, A. K. Y., Cheung, S. G., and Shin, P. K. S., 2017. Marine microalgae as dietary supplements in the cul- ture of juvenile Chinese horseshoe crabs,(Xiphosura)., 48: 3910-3924, DOI: 10.1111/are.13218.
Li, Z. J., Lin, L., and Yang, Y. Y., 2005. Effect ofcom- mercial probiotic on intestinal microflora of white shrmip,., 1 (3): 54- 59, DOI: 10.3969/j.issn.2095-0780.2005.03.009 (in Chinese with English abstract).
Liao, Y. Y., Hsieh, H. L., Xu, S. Q., and Kwan, B. K. Y., 2019. Wisdom of crowds reveals decline of Asian horseshoe crabs in Beibu Gulf, China., 53: 222-229, DOI: 10.1017/S003 060531700117X.
Looft, T., Allen, H., Cantarel, B., Levine, U. Y., Bayles, D. O., Alt, D. P.,, 2014. Bacteria, phages and pigs: The effects of in-feed antibiotics on the microbiome at different gut loca- tions., 8: 1566-1576, DOI: 10.1038/ismej. 2014.12.
Miao, F. Z., Zhao, Z. H., Li, Q. Z., Song, J., Wang, Y. J., and Hu, M. H., 2020. Impact of initial feeding and molting ongut microbiota., 77: 2847-2858, DOI: 10.1007/s00284-020-02108-x.
Nicholson, J. K., Holmes, E., Kinross, J., Burcelin, R., Gibson, G., Jia, W.,., 2012. Host-Gut microbiota metabolic inter- actions., 336: 1262-1267, DOI: 10.1126/science.1223 813.
Rognes, T., Flouri, T., Nichols, B., and Quince, C., 2016. VSEARCH: A versatile open source tool for metagenomics., 4: 2167-8359, DOI: 10.7717/peerj.2584.
Sekiguchi, K., and Nakamura, K., 1979. Ecology of the extant horseshoe crabs., 29: 37-45.
Shin, P. K. S., Li, H. Y., and Cheung, S. G., 2007. Horseshoe crabs in Hong Kong: Current population status and human exploita- tion. International Symposium on the Science and Conserva- tion of Horseshoe Crabs. Dowling Coll, Oakdale, NY, 347-351.
Shinohara, R., Sasaki, K., Inoue, J., and Hoshi, N., 2019. Butyryl- CoA:acetate CoA-transferase gene associated with the genusis decreased in the gut microbiota of Japanese pa- tients with ulcerative colitis., 38: 159-163, DOI: 10.12938/bmfh.18-029.
Shuster, C. N., 1979. Distribution of the American horseshoe ‘crab,’(L.)., 29: 3-26.
Sugita, H., Oshima, K., Tamura, M., and Deguchi, Y., 1983. Bac- terial flora of gastrointestine of freshwater fishes in river., 49: 1387-1395, DOI: 10.2331/suisan. 49.1387.
Sun, F. L., Wang, C. Z., Chen, L. J., Weng, G. Z., and Zheng, Z. P., 2020. The intestinal bacterial community of healthy and diseased animals and its association with the aquaculture en- vironment., 104: 775- 783, DOI: 10.1007/s00253-019-10236-z.
Tian, L., 2011. Re-discussion on probiotics., 8 (24): 251-252 (in Chinese with English abstract).
Wang, H., Li, Z. J., Li, C. H., Cao, Y. C., and Yang, Y. Y., 2009. Effects ofspp. on three common microalgae in shrimp ponds., 28 (2): 223- 227, DOI: 10.3969/j.issn.1000-8160.2009.02.012 (in Chinese with English abstract).
Wang, Y., Wang, Q. K., Xing, K. Z., Jiang, P., and Wang, J., 2021. Dietary cinnamaldehyde andimprove growth performance, digestive enzyme activity, and antioxidant capa- bility and shape intestinal microbiota in tongue sole,., 531: 75798, DOI: 10.1016/j. aquaculture.2020.735798.
Yang, B. B., and Shao, Q. J., 2013. Research progress of fish in- testinal microbes., 5 (23): 1-4, DOI: 10.3969/j.issn. 1004-3314.2013.23.001 (in Chinese with English abstract).
Yao, Y. Y., Yang, Y. L., and Gao, C. C., 2020. Surface display sys- tem for probiotics and its application in aquaculture., 12: 2333-2350, DOI: 10.1111/raq.12437.
Zeng, Y. H., Wu, N. C., and Chen, X. H., 2021.sp. nov. is an aerobic anoxygenic phototroph in the phylum Gemmatimonadetes., 11: 606612, DOI: 10.3389/fmicb.2020.606612.
J. Ocean Univ. China(Oceanic and Coastal Sea Research)
https://doi.org/10.1007/s11802-022-5205-x
ISSN 1672-5182, 2022 21 (3): 564-572
(September 23, 2021;
December 8, 2021;
January 15, 2022)
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2022
Corresponding author. E-mail: mhhu@shou.edu.cn
(Edited by Qiu Yantao)
 Journal of Ocean University of China2022年3期
Journal of Ocean University of China2022年3期
- Journal of Ocean University of China的其它文章
- Effect of Intertidal Elevation at Tsuyazaki Cove, Fukuoka,Japan on Survival Rate of Horseshoe Crab Tachypleus tridentatusEggs
- Asian Horseshoe Crab Bycatch in Intertidal Zones of the Northern Beibu Gulf: Suggestions for Conservation Management
- Experimental Investigation on the Interactions Between Dam-Break Flow and a Floating Box
- Variational Solution of Coral Reef Stability Due to Horizontal Wave Loading
- High Microplastic Contamination in Juvenile Tri-Spine Horseshoe Crabs: A Baseline Study of Nursery Habitats in Northern Beibu Gulf, China
- Influence of Autonomous Sailboat Dual-Wing Sail Interaction on Lift Coefficients