
棉属人工异源四倍体后代性状鉴定及花器转录组学分析
2024-01-22 06:44李昱樱荣二花吴玉香
作物学报 2024年2期
陈 天 李昱樱 荣二花 吴玉香
山西农业大学农学院, 山西晋中 030801
多倍体化在植物多样化、进化、驯化以及育种中发挥着极其重要的作用[1], 棉花是世界范围内经济价值最高的纤维作物, 同时也是研究多倍体进化的模式作物[2]。草棉(G.herbaceun, A1A1)是A 基因组二倍体栽培棉, 起源于非洲, 虽然没有大规模种植,但有着天然的优良性状, 如抗盐碱、抗旱、极早熟性等。雷蒙德氏棉(G.raimondii, D5D5)是D 基因组二倍体野生棉, 有着抗角斑病、生长旺盛、具有提高纤维细度和拉力的潜在能力。通过远缘杂交将亲本的有益性状结合起来, 能有效拓宽棉花种质资源,染色体加倍是恢复远缘杂种育性的最有效手段[3-4]。
分子标记技术基于DNA, 在植株整个生长期各个器官组织均可检测到, 不受环境影响; 多态性丰富; 数量多; 检测方便快捷等[5]。与其他的分子标记相比, SSR 是共显性标记, 能够提供更丰富的遗传信息, 已广泛用于杂种鉴定[6]、杂种纯度[7]及种质资源遗传多样性分析[8-10]中。
转录组(Transcriptome)是指特定生物体在某种状态或某一生理条件下, 细胞内所有基因转录产物的总和[11]。近年来本实验室进行了大量基于转录组测序在棉花育种上的应用与分析: 申状状等[12]对陆地棉和斯特提棉的杂种植株及人工异源六倍体植株(基因组AADDCC)进行无参转录组测序, 分析了三倍体杂种植株及异源六倍体植株与亲本的基因表达水平; 杨亚杰等[13]对草棉二倍体以及诱变获得的草棉三倍体和草棉四倍体进行转录组测序, 找到草棉多倍体在抗性及光合效率方面均优于二倍体的原因。随着棉花转录组测序研究的深入, 棉花基因组数据库逐渐完善, 为棉花新基因挖掘和种质创新提供了技术支持和数据库平台。
本研究以实验室前期人工合成的草棉与雷蒙德氏棉的人工异源四倍体(A1A1D5D5) S2、S3、S4代植株为材料, 并与陆地棉(G.hirsutum, AADD)进行纤维性状、生理生化、SSR 分子标记等方面的比较研究, 旨在将人工异源四倍体与自然界异源四倍体进行比较及性状鉴定。并通过花器转录组学分析, 比较人工异源四倍体与陆地棉的差异表达基因, 找到人工异源四倍体结实率低的真正原因, 为恢复其育性寻找有效途径, 进一步创新棉花种质资源。
1 材料与方法
1.1 试验材料
本研究试验材料采用实验室前期合成的草棉与雷蒙德氏棉的人工异源四倍体自交2 代(S2)、自交3代(S3)、自交4 代(S4)植株以及亲本和陆地棉TM-1。人工异源四倍体合成过程: 以栽培种红星草棉为母本和野生种雷蒙德氏棉杂交, 对杂种F1进行秋水仙碱诱变加倍[14]。异源四倍体植株逐年种植, 除去自然死亡的植株, 最后用于本研究共7 株, 分别用阿拉伯数字1~7 进行编号, 试验材料在培育前均进行65℃浸种4 h 及2%次氯酸钠消毒30 min, 苗期在人工气候箱培育, 待出现2 片真叶后移栽温室花盆,每穴1 株, 盆栽间隔0.5 m。草棉和陆地棉盆栽若干株, 分别取长势良好的3 株作重复。雷蒙德氏棉1株。研究材料具体信息见表1, 人工异源四倍体1~4号植株为S2代, 5~6 号植株为S3代, 7 号植株为S4代。多倍体后代株系关系如图1 所示, 其中2~5 号植株同期种植, 6~7 号植株同期种植。

表1 研究材料基本信息Table 1 Basic information of research materials
1.2 试验方法
1.2.1 生理指标测定 在试验材料整体进入现蕾期,选取不同植株新鲜嫩叶, 迅速用液氮预冷后转移至–80℃冰箱保存。根据北京盒子生工科技有限公司提供的试剂盒及方法分别测定丙二醛(malondialdehyde,MDA)含量、超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)活性。每个指标均设置3 个生物学重复, 使用96 孔板测定吸光值, 检测所用酶标仪型号为Multiskan FC。
1.2.2 SSR 分子标记 通过基于PCR 的简单重复序列标记 (simple sequence repeats, SSR)对提取的DNA 样品进行多态性分析。PCR 反应体系为10 μL:5 μL 2×TaqPCR Master Mix (含染料), 1 μL 正向引物(1 μmol L–1), 1 μL 反向引物(1 μmol L–1), 1 μL 模板DNA (50 ng μL–1), ddH2O 补齐。PCR 扩增程序: 94℃预变性5 min; 94℃变性60 s, 退火60 s, 72℃延伸90 s, 35 个循环; 72℃延伸10 min; 4℃保存。电泳程序及银染操作参照实验室前期方法[15], 显影至出现清晰条带后, 置于显影仪上人工读带, 有条带记1, 无条带记 0, 并计算引物的多态信息含量(polymorphism information content, PIC)。从棉花标记数据库(http://www.cottonmarker.org/Downloads.shtml)筛选11 对多态性好的SSR 引物, 信息见表2,由生物工程股份有限公司合成, 试验所用PCR 扩增仪型号为东胜ETC811, 电泳仪电源型号为北京六一DYY-6D, 电泳槽型号为北京六一DYCZ-24B, 显影仪型号为北京六一WD-9406。Marker 由北京索莱宝科技有限公司提供。PIC 计算公式[16]:

表2 SSR 引物序列信息Table 2 Sequence information of SSR primers
式中,k是一对SSR 引物扩增的等位位点的数目,Pi是第i个等位位点的频率。
1.2.3 转录组测序分析 由于无法准确判断杂种可育株的某个花器是否能成功结铃, 本研究选取S2代同期种植的2、3、4 号株(目前未结铃)的花器作为试验样品, 记为CL; 并以陆地棉TM-1 花器作为参照, 记为CK。以上试验样品均设置3 个生物学重复。将样品送百迈客生物科技有限公司进行RNA 提取及使用Illumina NovaSeq 6000 测序平台进行PE150模式测序。将下机原始数据(Raw Data)质控后获得的高质量序列(Clean Data)比对到陆地棉参考基因组上,进行后续分析。参考基因组的版本信息:Gossypium_hirsutum.TM_1.genome.fa。
1.2.4 数据处理 采用Microsoft Excel 2016 整理试验数据, 利用SPSS 26 软件进行数据处理与分析,利用Origin 2021 软件作图。
2 结果与分析
2.1 纤维性状观察
对人工异源四倍体及亲本纤维性状比较观察(图2)发现, 父本雷蒙德氏棉纤维呈褐色, 纤维粗短,母本草棉纤维呈白色, 人工异源四倍体纤维颜色介于亲本之间, 呈浅黄色, 纤维长于亲本。对人工异源四倍体和陆地棉纤维性状比较(图3)发现, 人工异源四倍体纤维长度与陆地棉接近, 但较陆地棉纤维稀疏, 浅黄色纤维性状较有优势。

图2 亲本及人工异源四倍体纤维形态Fig.2 Fiber morphology of G.herbaceum, G.raimondii, and artificial allotetraploid
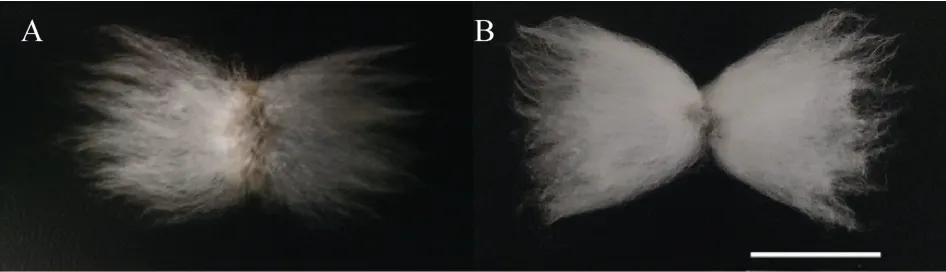
图3 人工异源四倍体及陆地棉纤维形态Fig.3 Fiber morphology of artificial allotetraploid and G.hirsutum
2.2 生理指标测定
研究材料由于遗传背景不同, 生理变化趋势也有所不同。对草棉、雷蒙德氏棉、人工异源四倍体后代以及陆地棉超氧化物歧化酶、过氧化物酶、过氧化氢酶活性和丙二醛含量进行检测发现(表3), 人工异源四倍体植株SOD、POD、CAT 活性及MDA含量均显著高于亲本和陆地棉, S2、S3、S4代之间酶活性有高有低, 无明显差异。表明人工异源四倍体在经过远缘杂交及加倍后, 抗逆性得到加强。
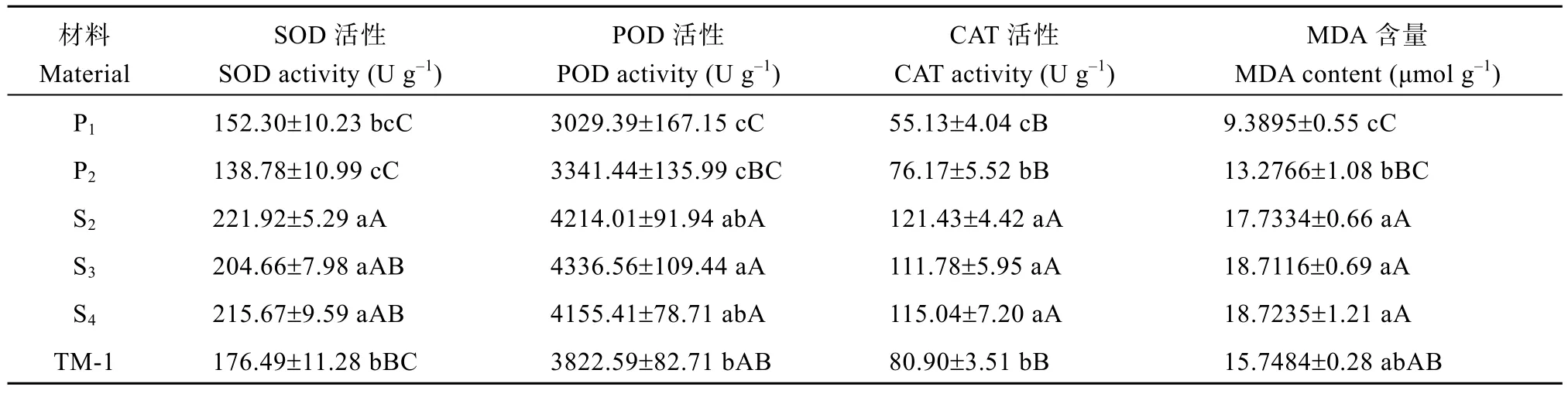
表3 各世代生理指标结果Table 3 Physiological indexes among different generations
2.3 SSR 分子标记
在棉花标记数据库筛选出11 对多态性好的SSR引物对7 株人工异源四倍体及亲本和陆地棉DNA 进行SSR 分子标记鉴定。SSR 扩增结果表明(表4), 11对引物总共扩增出59 个等位位点, 其中55 个位点具有多态性, 平均每对引物扩增出5 个多态位点。引物NAU2026 (图4-B)、引物NBRI_GL015 (图4-C)扩增出10 个位点数, NAU1169 扩增的位点数最少,扩增出2 个。11 对SSR 引物的多态信息含量PIC 在0.498~0.892 之间, 平均为0.742, 引物NAU2026、引物NBRI_GL015 的PIC 值最高, 说明这2 对引物多态性丰富, 能较好的揭示试验材料间的遗传多样性。引物NAU1169 的PIC 值最低。由图4 可知, 引物BNL4108、引物NAU2026、引物NBRI_GL015均可作为S2的特征引物, 引物NBRI_gM7 可作为人工异源四倍体的特征引物。

图4 部分SSR 引物扩增结果Fig.4 Part information of SSR primers
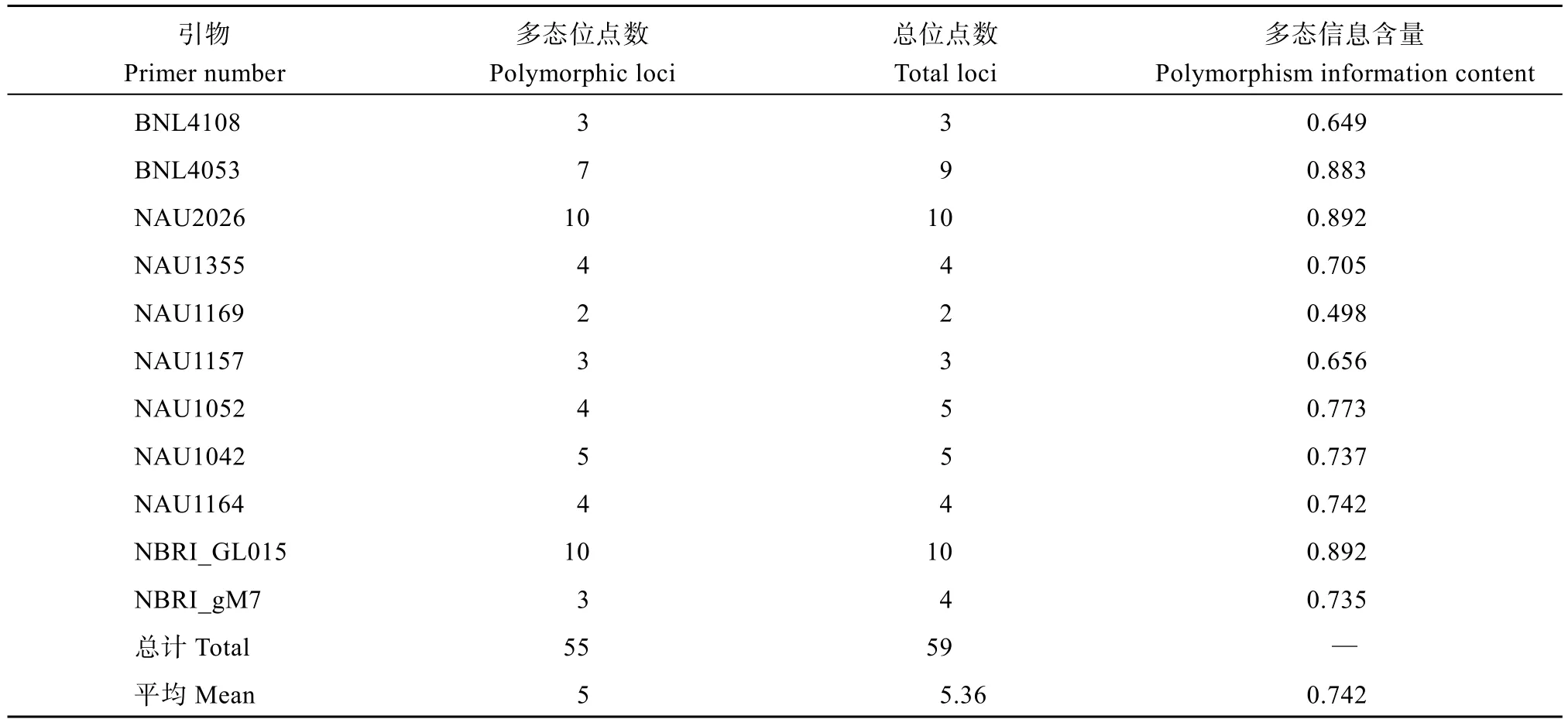
表4 SSR 引物扩增结果Table 4 SSR primers information
对人工异源四倍体及亲本的11 对SSR引物多态性位点进行统计分析(表5), S2代有13 个位点与母本相同, 有10 个位点与父本相同, 有28 个特异位点,平均每对引物有2.55 个特异位点, 有2 个缺失位点;S3代有9 个位点与母本相同, 有6 个位点与父本相同, 有7 个特异位点, 平均每对引物有0.64 个特异位点, 有12 个缺失位点; S4代有10 个位点与母本相同, 有6 个位点与父本相同, 有8 个特异位点, 平均每对引物有0.73 个特异位点, 有11 个缺失位点。分析表明人工异源四倍体后代的多态性位点在逐代减少, 推断是因为人工异源四倍体的基因片段随着繁殖代数增加在自行协调优化, 因此逐代自交能够提高人工异源四倍体育性。
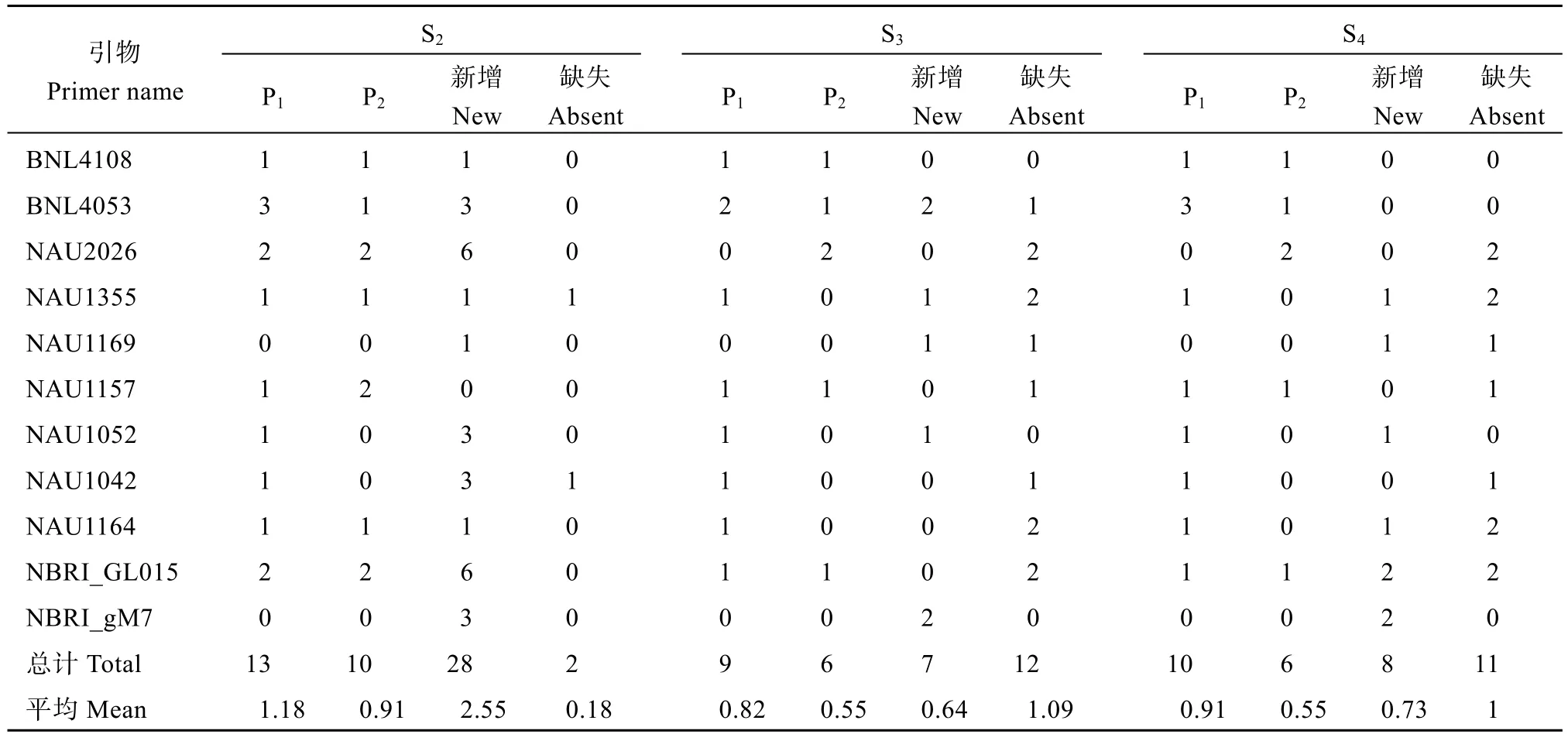
表5 人工异源四倍体及亲本SSR 引物多态性分析Table 5 Analysis of SSR primer polymorphisms for artificial allotetraploid and its parents
对人工异源四倍体相对于亲本扩增出的特异性位点进行分析(图5-A), S2的特异位点数最多,有28 条, S3、S4的特异条带数分别是7 条和8 条。其中各世代共同扩增出的特异条带有5 条, S2独有的特异条带有18 条, S3、S4均无单独扩增的特异条带。对人工异源四倍体与陆地棉的SSR 条带进行比较分析(图5-B), S2、S3、S4与陆地棉相同的条带分别是37 条、19 条、20 条, 其中有17 条是人工异源四倍体各世代与陆地棉共有的, 表明人工异源四倍体与陆地棉有相同的遗传背景, 其中 S2代同源性最强。
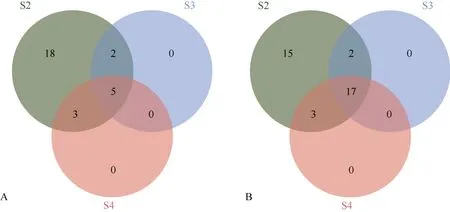
图5 人工异源四倍体的SSR 多态位点分布Fig.5 Resources of SSR polymorphism for artificial allotetraploid
2.4 转录组测序分析
2.4.1 差异表达基因的筛选 本研究转录组测序共获得38.29 Gb Clean Data, 各样品Clean Data 均达到6.08 Gb, Q30 碱基百分比在92.46%及以上, 分别将各样品的Clean Reads 与指定的参考基因组进行序列比对, 比对效率从94.42%到97.50%不等。采用FPKM 值对3 个生物学重复样品基因表达量进行标准化(FPKM > 1 的基因是表达的)。在陆地棉和人工异源四倍体的花器中分别有49,138 个和48,214 个基因表达, 其中有45,688 个基因是两者共同表达, 两者特异表达的基因分别是3450 个、2526 个。对陆地棉和人工异源四倍体花器中的表达基因进行比较,分析差异表达基因(differentially expressed genes,DEGs), 将差异倍数|log2(Fold Change) |≥2 且显著性FDR<0.01 作为筛选标准。在陆地棉与人工异源四倍体中一共筛选出5653 个差异表达基因, 其中上调表达基因有3062 个, 下调表达基因有2591 个。为了对差异表达基因分布有更直观地了解, 对其绘制了火山图(图6), 从图中也可以看出人工异源四倍体与陆地棉之间存在无差异表达的基因。

图6 差异表达基因火山图Fig.6 Volcanic map of differentially expressed genes
2.4.2 差异表达基因GO 富集分析 GO 注释系统包含生物学过程(biological process, BP)、细胞组分(cellular component, CC)和分子功能(molecular function, MF) 3 个主要分支。对筛选出的DEGs 进行GO富集分析, 结果表明, DEGs 共富集到3856 个GO 条目(term), 其中BP、CC、MF 分别富集到2422、520、914 个。对每个分类富集显著性Q-value 最小的前20个GO term 进行统计分析(图7), 在BP 中富集最显著的term 为“光合作用” (GO:0015979)、“蛋白质发色团连接” (GO:0018298)、“光合作用, 光吸收”(GO:0009765)、“对除草剂的响应”(GO:0009635)、“碳固定, 固碳作用” (GO:0015977)等; 在CC 中富集最显著的term 为“光系统II” (GO:0009523)、“光系统 I” (GO:0009522)、“光系统 I 反应中心” (GO:0009538)、“光系统II 放氧复合物” (GO:0009654)、“叶绿体类囊体膜” (GO:0009535)等; 在MF 中富集最显著的term 为“RNA 指导的DNA 聚合酶活性” (GO:0003964)、“ADP 结合” (GO:0043531)、“叶绿素结合” (GO:0016168)、“RNA-DNA 杂交核糖核酸酶活性” (GO:0004523)、“单氧酶活性” (GO:0004497)等。
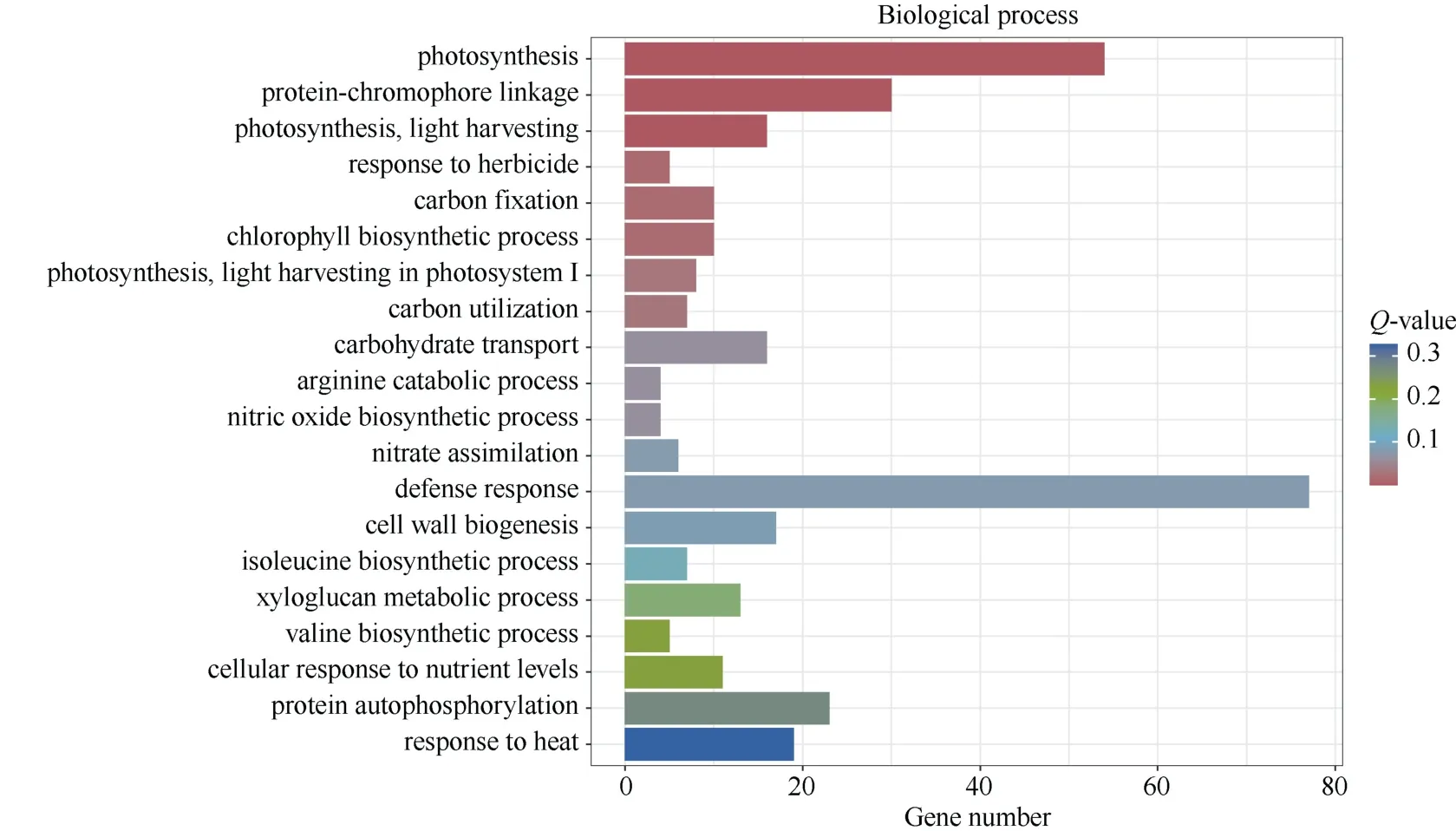
图7 差异表达基因GO 富集柱状图Fig.7 GO enrichment histogram of differentially expressed genes
2.4.3 差异表达基因KEGG 通路分析 对DEGs进行KEGG 富集分析, 对富集最显著的前20 个通路进行分析, 结果表明(图8), 人工异源四倍体与陆地棉DEGs 共富集到130 个通路, 其中最显著的通路是“光合作用-天线蛋白” (ko00196)、“光合作用”(ko00195)、“异黄酮的生物合成”(ko00943)、“苯并恶嗪类生物合成”(ko00402)、“2-氧代羧酸代谢”(ko01210), 富集的DEGs 数目分别是24、51、16、12、49 个。富集DEGs 数目最多的通路是“植物病原互作” (ko04626), 共富集到205 个DEGs。
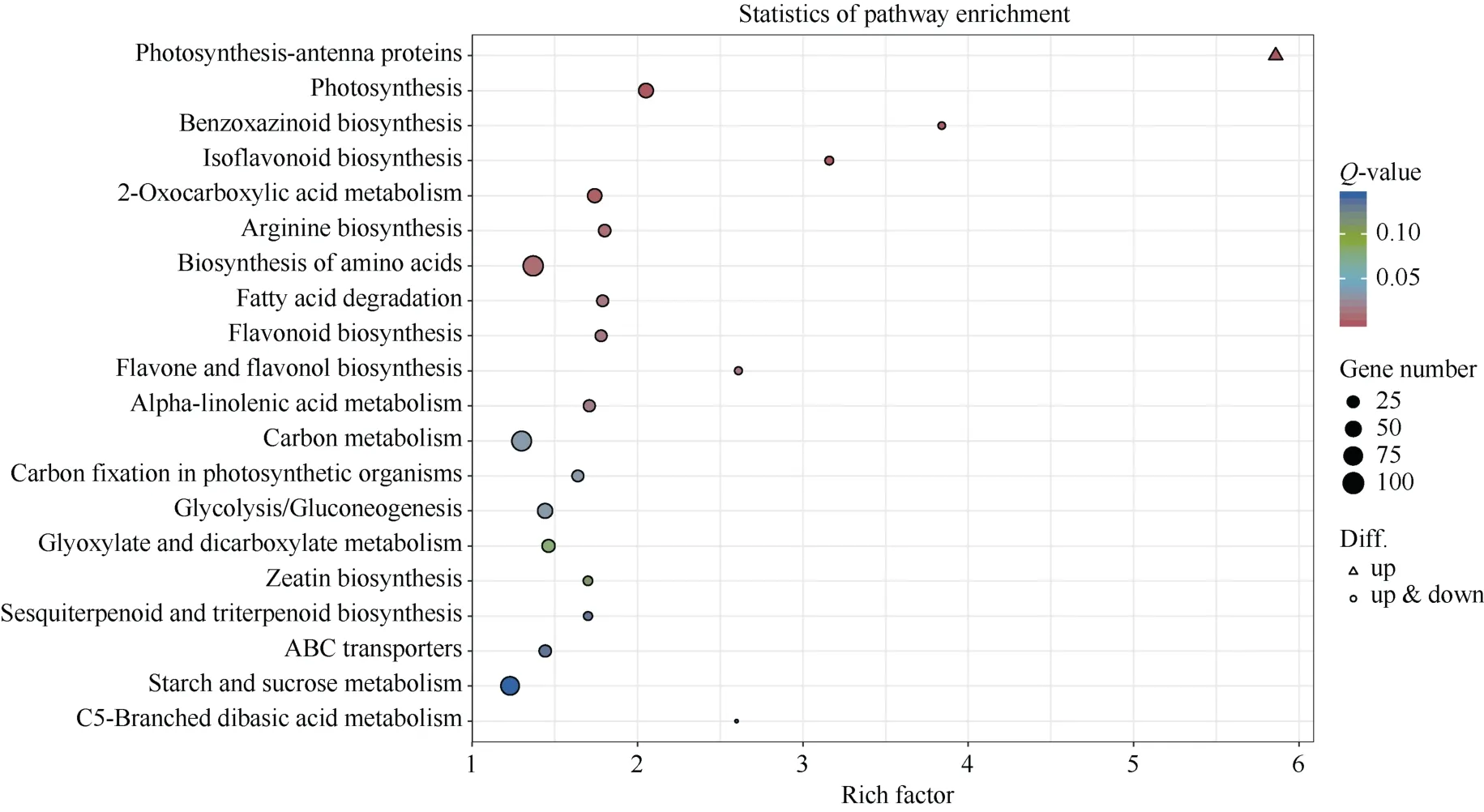
图8 差异表达基因KEGG 富集气泡图Fig.8 KEGG enrichment bubble diagram of differentially expressed genes
2.4.4 差异表达基因功能分类 植物通过光合作用为自身提供能量, 对植物的生长有重要意义。根据人工异源四倍体与陆地棉的差异表达基因KEGG富集最显著的前20 个通路分析结果(表6), 获得3个与光合作用相关的通路, 分别是“光合作用-天线蛋白” “光合作用” “光合生物中的碳固定”。在“光合作用-天线蛋白”通路中, 24个差异表达基因全部为上调表达。在“光合作用” “光合生物中的碳固定” 2个通路中上调表达的基因分别是70 个(93.33%)、25个(78.13%)。分析还表明一些与活性物质合成及代谢相关的基因也出现了差异表达, 其中与“氨基酸的生物合成”相关的基因有107 个, 且57 (53.27%)个上调表达; 有31 个基因与“类黄酮生物合成”相关, 其中22 个(70.97%)上调表达; 有32 个基因与“脂肪酸降解”相关, 其中22 个(68.75%)上调表达; 有55 个基因与“糖酵解/糖异生”相关, 其中42 个(76.36%)上调表达; 有89 个基因与“淀粉和蔗糖代谢”相关, 其中56 个(62.92%)上调表达。此外, 还有34 个基因与“ABC 转运蛋白”相关, 有18 个(52.92%)基因呈上调表达。
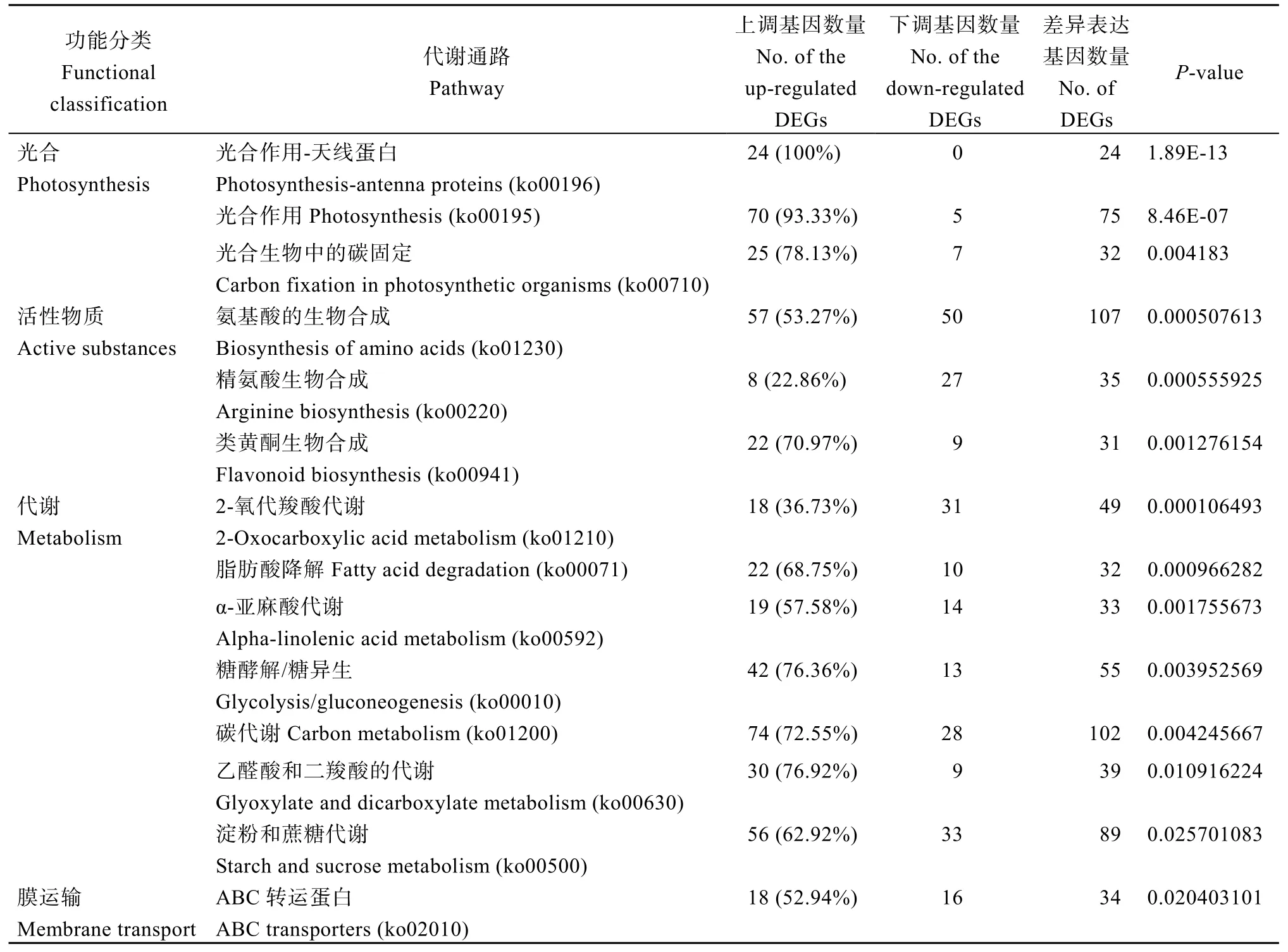
表6 CK vs CL 差异表达基因功能分类Table 6 Functional classification of differentially expressed genes in CK vs CL
与陆地棉相比, 人工异源四倍体除了“精氨酸生物合成” “2-氧代羧酸代谢”通路相关的基因上调表达少于下调表达外, 上调基因分别是8 个(22.86%)、18 个(36.73%), 其余的通路都是上调表达基因数量多于下调表达基因数量, 表明人工异源四倍体在光合、抗性、代谢等方面均表现出优势。
2.4.5 育性相关基因的筛选与分析 对可能影响人工异源四倍体育性的差异表达基因进行GO 功能注释, 结果如表7 所示, 共注释到23 个差异表达基因, 划分为7 个功能组, 这些功能组全部分布在生物过程类别中, 且注释到的下调基因数目多于上调基因数目, 下调基因17 个, 上调基因6 个。7 个功能组分别为“花粉萌发” “花粉识别” “花粉发育”花粉管发育” “花的发育” “花粉管导向” “花药发育”。其中“花粉萌发”注释到2 个差异基因, 1 个上调表达, 1个下调表达; “花粉识别”注释到的差异基因数最多,注释了14 个基因, 其中3 个上调表达, 11 个下调表达; “花粉发育”注释到3 个下调基因; “花粉管发育”注释到2 个下调基因; “花的发育”注释到2 个上调基因和1 个下调基因; “花粉管导向”注释到1 个下调基因; “花药发育”注释到1 个下调基因。

表7 CK vs CL 育性相关差异表达基因Table 7 Functional classification of differentially expressed genes in CK vs CL
利用在线工具NovoPro 对上述23 个差异表达基因进行蛋白信号肽预测, 对可能影响人工异源四倍体育性的差异表达基因进一步筛选。筛选结果如表8 所示, 总共筛选出 7 个候选基因, 分别是Gh_D04G029900、Gh_A01G062500、Gh_D02G100100、Gh_D01G067500、Gh_D05G168100、Gh_D02G075200、Gh_A05G248700, 7 个候选基因信号肽类型均为SP(Sec/SPI)。利用TMHMM2.0 在线软件对这7 个候选基因的蛋白跨膜结构域进行预测, 预测结果显示(表8), 7 个候选基因都有跨膜结构域, 其中差异基因Gh_A05G248700有3 个跨膜结构域, 其余6 个候选基因均有1 个跨膜结构域。利用在线工具Pfam 和TBtools软件对功能结构域预测, 预测结果显示(图9),7个候选基因均预测到4个相同的功能结构域, 胞外结构域是 D-甘露醇结合凝集素结构域(B_lectin,PF01453)、S 位点糖蛋白结构域(S_locus_glycop,PF00954), 胞内结构域是蛋白激酶结构域(Pkinase,PF00069)、蛋白质酪氨酸和丝/苏氨酸激酶结构域(PK_Tyr_Ser-Thr, PF07714), 其中Pkinase 结构域和PK_Tyr_Ser-Thr 结构域在候选基因的蛋白序列上有重叠。

图9 候选基因功能结构域Fig.9 Functional domain of candidate genes
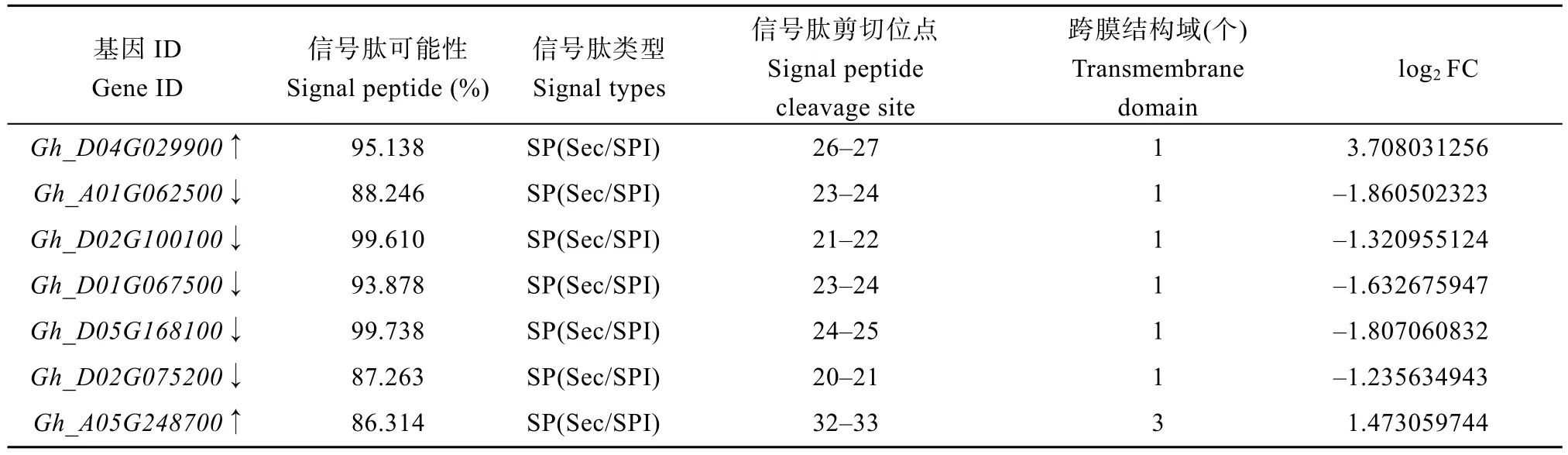
表8 候选基因信息Table 8 Candidate gene information
利用MEGA7 软件对候选基因进行聚类分析(图10)发现, 基因Gh_D01G067500与Gh_D05G168100亲缘关系最近, 与Gh_A01G062500聚为1 个小亚群, 其余4 个候选基因聚为1 个较大的群。由NR 注释列(表7)可知, 基因Gh_D01G067500、Gh_D05G168100、Gh_A01G062500均为G 型凝集素类受体丝氨酸/苏氨酸蛋白激酶, 且这3 个基因均下调表达, 因此这3个基因的差异表达可能是导致人工异源四倍体结实率低的主要原因。
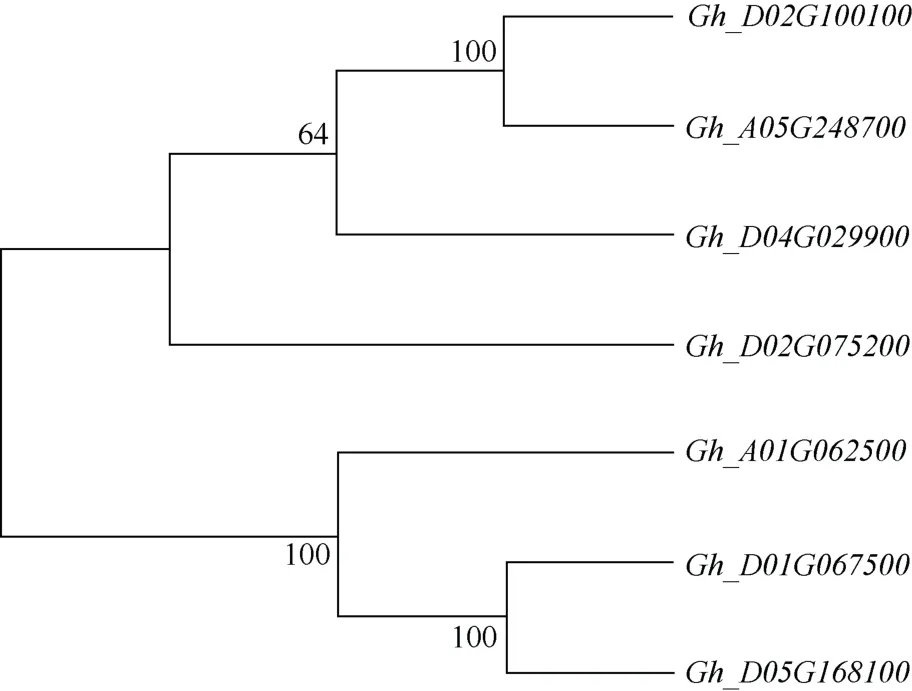
图10 候选基因聚类分析Fig.10 Cluster analysis of candidate genes
3 讨论
3.1 棉花远缘杂交及多倍体育种探讨
远缘杂交是人工合成新物种、新类型的重要手段, 在远缘杂交的后代中, 会出现因自然选择而消失了的中间物种或类型, 同时也会打破种间限制,把2 个或多个物种经过自然界长期进化积累的有益特性重新组合, 形成新的类型或新种。但是亲缘关系较远的种间杂交时, 得到的杂种是高度不育的,为打破这种杂种后生殖隔离, 使用秋水仙碱对杂种进行染色体加倍是恢复育性的有效手段。实验室前期Wu 等[14]研究发现, 草棉与雷蒙德氏棉二倍体杂种后代的花粉母细胞存在大量的异常减数分裂行为,导致杂种高度不育。但对二倍体杂种加倍后, 所获得的人工异源四倍体早代出现结铃, 表明育性有所恢复。
棉花通过远缘杂交加倍获得新种有许多成功的例子, 聂以春等[17]1985 年以培育的亚洲棉品种与野生种司笃克氏棉杂交获得杂种, 并加倍成异源四倍体, 经过多年自交培育出植株形态和农艺性状稳定、结铃性强的异源四倍体(亚洲棉×司笃克氏棉)。目前关于草棉与雷蒙德氏棉的人工异源四倍体的研究鲜有报道, 本研究以人工异源四倍体为材料, 从生理指标结果可以看出, 在温室生长环境中(生长期温度控制不高于35℃), 其SOD、POD、CAT较亲本及陆地棉活性均显著增加, MDA 含量提高,表明其抗逆性较亲本增强, 可能的原因是多倍体植株同一位点基因剂量加倍, 从而导致基因表达量增强, 引起植株表型和生理状态发生改变。Jiang 等[18]使用定量遗传方法QTL 分析陆地棉与海岛棉F2后代中的纤维相关性状, 发现影响纤维产量和质量的基因座大多数是在D 基因组而不是在A 基因组中,这解释了相比较于二倍体栽培棉, 异源四倍体栽培棉纤维产量和品质的优越性。本研究的异源四倍体纤维长度比亲本长, 与陆地棉接近; 并且纤维颜色为彩色, 介于亲本之间, 具有彩色纤维相关基因的挖掘潜力, 可以作为纤维性状遗传的研究材料, 后期可以与陆地棉或海岛棉杂交, 有望育成天然彩棉新品系。
自然界棉属异源四倍体祖先供体种是棉属进化研究的热点。多年来普遍一致的看法是雷蒙德氏棉是D 基因组的祖先供体种[19], 但A 基因组的祖先供体种一直存在争议。Huang 等[20]最近采用测序及Hi-C 技术第一次组装了非洲草棉A1基因组, 并重新组装了高质量已测序的亚洲棉石系亚A2和陆地棉标准系TM-1 基因组, 揭示了所有现存的A 组都可能起源于共同的祖先A0, 而不是现存的二倍体A1和A2。本研究人工异源四倍体与陆地棉基因组相同,SSR 分子标记显示, 人工异源四倍体不仅扩增出亲本的互补条带与特异性条带, 还扩增出与陆地棉相同的条带, 这为研究棉属系统进化提供了有价值的材料。同时SSR 标记特异性分析显示, 随着人工异源四倍体自交繁殖代数增加, 条带特异性呈现逐渐降低的趋势, 这与盖树鹏的研究结果一致[21]。
3.2 棉花转录组研究探讨
转录组研究是基因功能及结构研究的基础和出发点, 已广泛应用到棉花生长发育、非生物胁迫、突变表型和分子育种等领域。陈全家[22]对陆地棉和海岛棉2 个棉种胚珠进行转录本测序, 筛选差异基因并分析, 认为GbMYB25与GbDET2基因可能参与调控棉花纤维起始发育。邵冰欣[23]发现基因GbDRP66319和GbHIR42734沉默表达的棉花植株比对照株黄萎病发病更严重, 认为基因GbDRP66319和GbHIR42734在棉花抗黄萎病方面起作用。包秋娟[24]对早熟棉种“新陆早17”进行干旱胁迫处理, 发现处理之后的棉花植株可诱导油质蛋白基因GhOleosin1的表达。本研究对人工合成的草棉与雷蒙德氏棉人工异源四倍体与陆地棉进行转录组测序,共获得5653 个差异基因, 其中上调表达基因有3062个, 下调表达基因有2591 个, GO 富集分析显示DEGs 共富集到3856 个GO 条目, 在光合作用、光系统II、RNA 指导的DNA 聚合酶活性所占比例较高, 对差异表达基因KEGG 富集表明, 差异基因共富集到130 个代谢通路, 主要富集在光合作用-天线蛋白、光合作用和异黄酮的生物合成相关代谢通路等, 表明人工异源四倍体在光合作用和抗逆等方面有着多倍体及杂种优势。此外, 为了探究人工异源四倍体结实率低的关键因素, 我们进一步对差异基因进行了筛选, 总共筛选出7 个影响人工异源四倍体不育的候选基因, 发现这些基因影响花粉发育及识别, 最终导致花粉败育, 这符合我们观察发现的人工异源四倍体花粉出现干瘪的现象。
3.3 植物凝集素类受体蛋白激酶功能探讨
凝集素是植物生命的基础, 在细胞信号识别、细胞发育、细胞防御有着重要作用[25]。早期人们对凝集素类受体蛋白激酶家族的研究停留在基础结构研究和基因表达层面, 随着生物信息技术的发展,一系列研究表明凝集素表现出不同的分子机制, 如花粉发育、雄性不育、抗病、应激反应、激素信号传导、植物-真菌共生等。Walker 等[26]在玉米中鉴定出了第一个G 型凝集素类受体蛋白激酶ZmPK1, 该蛋白激酶的催化结构域通过跨膜结构域连接到一个类似于芸薹属自交不亲和位点中编码的糖蛋白的胞外结构域, 这些糖蛋白参与花粉和柱头之间的识别系统。Zuo 等[27]通过cDNA 文库筛选, 从陆地棉中克隆了一个凝集素类蛋白激酶新基因GhlecRk, 并对其进行表达模式分析得出GhLecRK在发育中的棉铃富集, 在根和茎中含量较低, 在叶中没有表达。结构域分析显示GhlecRK蛋白具有多个结构域, 包括ATP 结合位点、一个跨膜区、一个凝集素类结构域和一个激酶结构域, 判断GhlecRK是一种凝集素类蛋白, 在纤维发育阶段参与调控。对本研究中筛选的育性候选基因Gh_D01G067500、Gh_D05G168100、Gh_A01G062500进行结构域分析,这3 个候选基因的胞外都含有S-locus 结构域, 此结构域在被子植物自交不亲和系统中起作用[28]。Wan等[29]研究表明, 一个凝集素类受体蛋白激酶基因LecRK-IV.2突变引起花粉发育缺陷导致拟南芥雄性不育, 该突变表型可能是由该突变引起的一种尚未明确的孢子体缺陷引起的, 因此LecRK-IV.2在花粉发育中起关键作用。毕真真[30]在水稻中发现一个凝集素类受体激酶基因OsL-LecRK7, 通过半定量RT-PCR 说明OsL-LecRK7调控花粉发育, 随之将OsL-LecRK7突变体与野生型正反交发现结实率显著提高。因此后续期望将人工异源四倍体与陆地棉进行正反交试验来克服自交不亲和性, 进而恢复其育性。
4 结论
新合成的草棉与雷蒙德氏棉人工异源四倍体的纤维颜色呈浅黄色, 长度与陆地棉接近。人工异源四倍体SOD、POD、CAT 活性以及丙二醛含量均显著高于亲本及陆地棉。SSR 分子标记鉴定结果表明,S2、S3、S4与陆地棉相同的条带分别是37 条、19 条、20 条, 其中17 条是各世代与陆地棉共有。转录组测序结果表明, 人工异源四倍体与陆地棉中共有5653个差异表达基因; 富集发现人工异源四倍体在光合和抗逆方面均优于陆地棉; 通过对育性相关基因聚类, 基因Gh_D01G067500、Gh_D05G168100、Gh_A01G062500均为G 型凝集素类受体丝氨酸/苏氨酸蛋白激酶, 这可能是导致人工异源四倍体自交不亲和的关键基因。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
海洋通报(2022年4期)2022-10-10
农业工程学报(2022年5期)2022-06-22
长江蔬菜(2021年12期)2021-04-04
中华戏曲(2020年2期)2020-02-12
中国果业信息(2019年11期)2019-01-05
现代园艺(2017年13期)2018-01-19
中成药(2016年4期)2016-05-17
西南农业学报(2016年5期)2016-05-17
华东理工大学学报(自然科学版)(2015年3期)2015-11-07
