
谷子中矮秆资源创制、遗传解析和育种利用
2024-01-22 06:43刁现民王立伟李顺国程汝宏
作物学报 2024年2期
刁现民 王立伟 智 慧 张 俊 李顺国 程汝宏
1 作物基因资源与育种全国重点实验室 / 中国农业科学院作物科学研究所, 北京 100081; 2 河北省农林科学院谷子研究所, 河北石家庄 050035
谷子是中国起源的特色作物, 栽培历史悠久,是中华农耕文明的载体作物。谷子抗旱节水特性突出, 是旱作生态农业可持续发展的主栽作物, 在我国北方粮食安全和产业振兴中具有重要地位[1]。谷子生产对农业多样性至关重要, 西北地区和东北地区水资源和旱情日益加重, 华北地区地下水超采严重形成巨大的地下漏斗, 抗旱节水的谷子成为首选的替代作物, 展示出谷子生产的广阔前景, 但倒伏是限制谷子机械化和产业化水平提升的关键因素之一。株高是作物的核心株型性状和控制产量的主要性状, 也是解决倒伏问题和适应机械化作业的关键性状。20 世纪60 到70 年代, 小麦和水稻等主要农作物的绿色革命, 基本解决了倒伏和耐肥水问题,带来了产量的显著提升, 同时株高的降低, 也为后来的机械化收获提供了方便[2-5]。对小麦和水稻绿色革命基因Rht和SD1的遗传和分子解析, 并结合其他育种技术的提升, 使作物抗倒伏和株型育种提高到了一个新高度, 也带来小麦水稻生产潜力持续攀升[6], 同时, 水稻和小麦在矮秆抗倒伏育种上的成功也为其他作物提供了经验。
虽然谷子是我国起源的古老作物, 但遗传育种水平和水稻小麦差距巨大, 到20 世纪90 年代中后期, 谷子生产用品种仍以高秆为主, 夏谷一般株高130~160 cm, 春谷150~170 cm, 个别品种的株高甚至超过200 cm, 倒伏经常发生, 更不适合机械化田间管理和收获。因此, 进行中矮秆谷子亲本创新和育种成为生产的迫切需要。我国广大谷子科技工作者很早就重视中矮秆谷子资源的创制和利用, 先后培育出延矮、安矮、郑矮号等各类矮秆材料, 但一直缺乏对矮秆材料的系统整理, 矮秆材料在育种和生产中的利用有限。20 世纪90 年代, 矮88 的育成和吨谷在育种和生产上的利用标志着谷子中矮秆利用的新时代。杜瑞恒等[7]1995 年对我国谷子矮秆材料曾做过综述性总结, 但近年来谷子矮秆材料的创制、遗传分析和育种应用都发生了很大变化, 本文对谷子矮秆材料的创制和育种利用进行系统的梳理和综述, 以期为谷子育种和生产利用提供综合信息。
1 谷子中矮秆资源创制及其类型
1.1 谷子矮秆材料的创制
1961 年印度学者Ratnaswamy 和Dhanaraj 报道,从栽培谷子品种Co2 中发现一个株高降低50%且不倒伏的矮秆材料, 这是首个矮秆谷子的报道[8]。早在20 世纪70 年代, 我国谷子科技工作者就开始了各类谷子矮秆材料的创制和选育, 以陕西省延安地区农业科学研究所利用辐射诱变获得的延矮1 号为最早, 但正式报道要到1986 年师公贤在《陕西农业科学》发表矮秆谷子遗传分析[9]。谷子的矮秆材料来源包括辐射诱变、EMS 诱变、自然变异和远缘杂交等, 以辐射诱变的居多。河南省农业科学院作物科学研究所丁如坤[10]利用Co60处理农家品种郸城六茬白, 处理当代和新农724 杂交, 在后代选育出了73r19、78r07 和郑71 矮等矮秆品系。孟昭桂等[11]利用Co60处理夏谷品种青到老, 从处理后代选育出郑矮2 号等矮秆材料。除物理诱变外, 化学诱变也是创制矮秆材料的有效方法, 中国农业科学院作物科学研究所刁现民团队利用甲基磺酸乙酯(EMS)诱变获得了多个谷子矮秆材料[12-14]; 山西农业大学袁蕊[15]也成功利用EMS 诱变创制了谷子矮秆材料。在自然变异利用方面, 李东辉在夏谷品种宁黄1 号大田发现了矮秆突变体, 自交稳定后培育成矮宁黄新品种。姚占廷在丰产品种昭谷1 号大田发现了矮秆突变体, 自交稳定后获得了84133 矮秆纯系, 并利用这个材料培育出蒙古6 号矮秆品种[16]。印度学者Ratnaswamy 和 Dhanaraj[8]报道的 Co2 矮秆和Dineshkumar 等[17]报道的3 个矮秆sic24、sic25 和sic26 均是自然产生的突变体。谷子品种资源中很多矮秆材料来源于自然变异, 如农家品种的大青秸和宽九等; 还有很多矮秆材料来源不清, 这样的材料多数是自然变异。在远缘杂交产生矮秆方面, 河北省沧州地区农业科学研究所朱秀华用谷子和高粱杂交, 尽管其机制尚不清楚, 但也产生了各类矮秆变异, 特别是叶穗直立的矮秆材料已在育种中得到应用[18]。矮协1 号和矮协2 号等矮秆材料也是通过不同地理类型间谷子杂交, 在后代中选育出来的。杜贵和崔文生等利用谷子高度雄性不育系进行杂交或构建“动态基因库”, 选育出坝矮号矮秆谷子材料[19]。尽管广大谷子科技工作者在育种实践中创制了多种多样的谷子矮秆材料, 但对矮秆材料系统梳理相对缺乏, 杜瑞恒等[7]报道过对谷子矮秆材料的梳理和GA 敏感性测定, 以后刁现民团队收集了各类谷子矮秆材料, 测定了这些材料矮秆性状的显隐性[20], 结合文献报道, 将谷子矮秆材料整理如表1所示。

表1 谷子矮秆材料的显隐性Table 1 Dominant or recessive of dwarf foxtail millet
1.2 谷子矮秆材料对GA 敏感性和类型
作物矮秆材料的形成多和赤霉素(GA)的合成及信号传导通路的基因突变有关。对GA 敏感的材料多数是GA 合成途径基因的改变, 而对GA 不敏感的材料多是GA 受体或信号通路基因的突变。杜瑞恒等[7]试验表明, 我国多数谷子矮秆材料对GA 敏感,如矮宁黄、安矮5 号、麦谷1 号、矮协1 号等, 仅赤峰市农业科学研究所发现的84133 为不敏感。钱继岳等[20]对近50 份谷子矮秆进行的鉴定, 也发现只有84133 对GA 不敏感。陈金桂等[21-22]对84133、623C和矮宁黄进行GA 处理, 也发现84133 不敏感, 而623C 和矮宁黄对GA 敏感, 外施GA 可以恢复623C和矮宁黄的株高。赵丽娟等[23]对谷子辐射诱变形成的矮秆突变体d93090 的分析, 表明其也是GA 敏感型。印度学者Dineshkumar 等[17]报道的3 个谷子矮秆材料对GA 不敏感。从这些结果看出, 谷子的矮秆材料多数是GA 合成过程中的变异, 少数是GA 信号通路或者其他机制导致的变异(表1)。除GA 外, 一些谷子矮秆可能是油菜素内酯(BR)系统发生的变异,如T539[24]。
谷子矮秆材料划分为穗下垂型和穗直立型两类,这主要是基于形态上的表现。矮宁黄、吨谷、宽九、济矮5 号等矮秆材料的株型和高秆材料没有显著改变, 苗期叶片较宽且深绿, 拔节后叶片披垂, 抽穗后穗下弯。这种类型属于下垂型, 此类材料株高的降低多数是由于节间的缩短造成, 植株的节数和叶片数没有改变。矮协1 号、延四直、矮88、晋汾矮6 号等矮秆材料在植株变矮的同时, 叶片变短且直立, 穗颈变短穗直立, 整体表现了紧凑的株型。无论是下垂型还是直立型矮秆, 由于株高的降低, 抗倒伏性均显著提高。下垂型矮秆由于叶片大小和形态未改变, 仍存在着株间遮挡郁闭的问题, 耐密性较差; 直立型矮秆整体株型紧凑, 单株占据空间小,相对耐密植。但这类材料多数在降低株高的同时,伴随着穗变小、秕谷率高等众多问题。
1.3 谷子矮秆材料生理特征
多年多点对谷子矮秆材料的观察和试验发现,所有矮秆材料在株高降低的同时, 也表现了明显的早衰, 主要特征是后期生长势弱, 籽粒灌浆受影响,秕谷率高, 影响产量的提高。籍贵苏等[25]对高秆和矮秆谷子品种的根系进行了比较, 高秆组的品种包括181、8337、宁229、高39、青丰谷和豫谷1 号, 株高在125 cm 左右, 叶片披散, 穗于顶部弯曲下垂;矮秆组材料包括矮88、麦谷1 号、矮宁黄、辽水、737 青、交176 等, 株高在60~80 cm, 有披散和紧凑2 种株型。分析发现矮秆材料根系量显著低于高秆材料, 说明根系弱可能是矮秆材料早衰的原因。通过对高矮秆品种个体发育与产量的相关分析, 矮秆比高秆品种前期发育早, 但植体灌浆期干物质积累和转移变化不明显; 高秆品种植株灌浆期干物质积累和转移变化明显, 籽粒产量源于植株中的营养转移和开花后的光合产物积累[26]。
印度学者Dineshkumar 等[17]报道的sic 24、sic 25和sic 26 三个矮秆材料表型完全相似, 开花期株高为73.53 cm, 而高秆对照为127.45 cm; 矮秆材料的分蘖数平均为7.62 个, 高秆对照5.28 个; 高秆和矮秆材料的节数是一样的, 差别在节间长度。就单株生物量而言, 矮秆低于高秆材料, 但矮秆的收获指数(the harvest index, HI)为 41.43%, 高于高秆的38.64%。
袁蕊[15]对EMS 处理晋谷21 获得的矮秆材料的分析发现, 矮秆材料叶片中的叶绿素a、叶绿素总量和类胡萝卜素含量低于晋谷21, 叶绿素b含量只有在抽穗期高于晋谷21, 其他生育期均低于晋谷21;拔节和抽穗期矮秆突变体叶片中的可溶性总糖含量均低于晋谷21, 灌浆期却高于晋谷21, 还原糖和纤维素在各生育期都低于晋谷21, 淀粉含量在拔节和灌浆期均高于晋谷21; 矮秆材料茎粗和分蘖数显著高于晋谷21, 穗长和总节数没有显著变化, 每段节间长极显著或显著低于晋谷21, 叶宽和叶面积显著高于晋谷21, 但是其穗重、粒重、茎叶重和总生物量方面显著低于晋谷21, 千粒重没有显著变化。
2 谷子矮秆性状的遗传解析和基因定位
2.1 谷子矮秆材料的遗传控制
目前发现的谷子矮秆材料除少数几个外, 多数未进行遗传分析。师公贤等[9]将矮秆材料延096 与高秆不育材料安A 进行杂交, 证明延096 的矮化性状由隐性单基因控制。延096 是由Co60辐射诱变而来, 所以当时认为经过诱变产生的谷子矮秆突变体都是隐性单基因遗传突变造成的。孟昭桂等[11]将株高70 cm 的郑矮2 号和株高120 cm 左右的高秆材料衡研130 和青到老杂交, 2 个F1代均表现高秆, 说明郑矮2 号的矮秆为隐性。郑矮2 号和衡130 的F2表现15∶1 的分离比例, 说明以衡研130 为背景, 郑矮2 号含有2 个隐性矮秆基因。但郑矮2 号和其原始来源品种青到老杂交的F2, 表现了3∶1 的分离比例,说明在青到老背景下, 郑矮2 号的矮秆基因为单基因隐性。也意味着青到老自身含有一个衡研130 不具备的矮秆基因。郑矮2 号和来自陕西省农业科学院粮食作物研究所的株高为68 cm 的7516 杂交, F1代表现高秆, 说明郑矮2 号和7516 分别带有不同矮化基因。Qian 等[20]通过高矮秆材料杂交, 根据F1植株的表现判断矮秆基因的显隐性, 结果发现我国的矮秆材料除赤峰的84133 外, 其他都是隐性的, 这和84133 的发现者姚占廷等[16]的分析结果基本一致。Dineshkumar 等[17]发现的sic 24、sic 25 和sic 26表现为显性遗传, 这3 个矮秆材料是一个来源, 很可能是相同的矮秆基因。综合所有这些结果可以看出, 目前发现的谷子矮秆材料, 以隐性为主, 显性矮秆材料很少。从形态上看, 隐性矮秆表现了较高的多样性, 而显性矮秆均表现为粗壮宽叶, 相对单一的表型。
通过矮秆材料间杂交, 根据F1表型来判断父母本所具有的2 个矮秆基因的等位性, 在不同作物矮秆材料遗传分析中都采用这种方法, 但谷子这方面的工作相对少。孟昭桂等[11]通过郑矮2 号和7516 杂交, 判断出郑矮2 号和7516 分别带有不同矮化基因;高俊华等[27]通过安矮3 号和延矮1 号的杂交, 发现F1为矮秆, 推断安矮3 号和延矮1 号带有等位的矮秆基因。Qian 等[20]也进行了大量的矮秆材料间杂交,配制杂交组合45 个, 发现赤矮9 号、1066A 和矮丰1 号含有等位的隐性矮秆基因, 宽九和矮协1 号带有等位的隐性矮秆基因; 而多数矮秆材料间所携带的矮秆基因是不同的, 说明谷子矮秆基因的丰富多样性。总体来说, 谷子矮秆材料这方面的研究还很初步, 有关的结果还需要核实和验证, 这对认识不同矮秆材料所携带的矮秆基因是否相同有重要意义。
2.2 谷子矮秆相关基因定位
谷子最早的基因定位系统是河北省农林科学院谷子研究所王润奇等[28]创制的三体定位系统, 利用这个系统高俊华等[27]将安矮3 号的矮秆基因定位到谷子3 号染色体上, 尽管没有定位区间, 但在没有分子标记的时代是最先进的研究体系, 他们利用这个体系还进行了其他农艺性状的定位, 并进行了这些性状间的连锁关系分析[29]。随着分子生物学的快速发展, 谷子分子标记开发也取得长足进步, 为谷子矮秆基因的精细定位和克隆奠定了基础。中国农业科学院作物科学研究所谷子基因资源团队, 率先利用分子标记对多个谷子矮秆材料进行基因定位和克隆。延4 直是陕西省延安地区农业科学研究所发现的一个叶片和穗直立的矮秆突变体, 延4 直和豫谷1 号杂交, F1表现为野生型高秆表型, F2表现高秆195 株和矮秆64 株的分离, 符合隐性单基因控制的分离比例。利用BSA 混池法, 首先将该矮秆基因定位到5 号染色体标记P17X 附近, 再开发标记进行精细定位, 最后将目标基因定位到 YGY5045 与YGY5082 两个标记之间254 kb 的物理距离区间[30]。谷子体细胞变异也是矮秆材料的一个来源, T22 和T25 都是豫谷1 号组织培养后代中发现的矮秆材料,这2 个材料与野生型豫谷1 号相比在株高、节长、穗长和根系等方面都有明显的缩短现象。通过石蜡切片显微结构观察表明, 突变体茎部细胞长度明显减小、宽度明显增加。T22 和T25 的植株高度能够通过喷施赤霉素来恢复正常水平, 属于赤霉素敏感型突变体。将 T25 突变体与正常株高谷子品种SSR41 杂交, F1表现出正常株高表型, F2共获得2560株, 其中615 株为矮秆, 1945 株正常株高, 经χ2检测正常株与矮秆株符合3∶1 分离比, 因此认为该突变体矮化表型是由一个隐性单基因控制。通过对F2群体隐性单株采用BSA 混池法进行初定位, 找到了与目标基因较紧密连锁的分子标记b215 和b198, 再经过精细定位, 最终定位到了3 号染色体标记fxj032和fxj037 之间52.7 kb 的区间内。根据豫谷1 号全基因组序列, 预测在所定位区间的候选基因, 发现共有12 个完整的开放阅读框, 对此12 个基因进行表达量分析, 赤霉素合成关键酶GA20ox-2基因在突变体中表达量比野生型明显升高, 表明T25 突变体可能是GA 合成途径的相关基因发生了变异[13]。谷子矮秆小穗突变体si-dw3是从豫谷1 号EMS 诱变库中筛选得到的, 突变体茎缩短变细, 节数减少, 节间长度均缩短; 穗变小, 一级分枝数量减少, 种子长度增加; 叶缩短, 叶数目减少。通过茎的组织切片分析, 发现茎细胞长度缩短。si-dw3对外源赤霉素的响应与野生型一致, 但其株高不能完全恢复。以si-dw3为母本, 具有较高株高的晋汾8 号为父本构建作图群体, 进行遗传分析和基因定位。结果表明si-dw3的突变性状由隐性单基因控制。利用图位克隆和分子标记技术将突变基因定位在 8 号染色体标记SNP10-1 和SNP7-1 之间296.7 kb 的范围内[14]。
谷子上第一个克隆和进行功能验证的矮秆基因是对84133 矮化基因的功能分析(图1)。利用84133和张矮10 号的RIL 群体F6剩余杂合体、以及84133和豫谷1 号杂交的F2群体, 分析发现84133 和张矮10 号的剩余杂合体群体(Residual heterozygous line,RHL)表现了70~80 cm∶40~50 cm∶20~30 cm 株高植株1∶2∶1 的分离比例; 而84133 和豫谷1 号的F2群体也表现了120~30 cm∶65~75 cm∶30~40 cm植株1∶2∶1 的分离比例, 表明84133 含有一个半显性矮秆基因。利用图位克隆的方法将该基因定在9 号染色体标记Si332 和Si188 之间72 kb 的基因组区段, 该区域有 6 个转录本编码基因, 其中编码DELLA 蛋白的基因SiDw1被认定为目标基因。该基因与拟南芥的GAI/RGA, 水稻的SLR1, 玉米的D8,小麦的Rht-B1b和Rht-D1b具有很高的同源性, 属于GRAS 家族的DELLA 蛋白基因。序列分析表明, 在84133 矮秆突变体中SiDw1的编码区被一个约5.5 kb的copia反转座子插入, 从而造成转录本被截断, 形成一个缺失 N 端的截短转录本(SiDw1-TT); 同时,还形成一个DELLA 蛋白N 端与copia的TLR 的融合转录本(SiDw1-CT), 而原始野生型转录本为SiDw1-WT[31]。Southern 杂交和 RACE 分析发现84133 基因组中还含有一个未被copia插入的正常SiDw1基因, 这可能是copia插入过程造成的基因复制。将SiDw1-TT在豫谷1 号谷子和水稻中超表达,谷子和水稻的转基因植株均表现了明显的矮秆, 表型和84133 一致, 说明是SiDw1-TT导致的矮秆。这是谷子上克隆的第一个并进行了转基因验证的功能基因。

图1 谷子半显性矮秆基因DWARF1 的图位克隆Fig.1 Map-based cloning of foxtail millet semi-dwarf gene DWARF1 (Zhao et al.[31])
2.3 谷子株高遗传的QTL 分析
典型的矮秆材料很容易辨别, 遗传分析也相对容易。但在谷子自然群体中, 株高变异从几十厘米到数米, 说明谷子资源群体中存在着丰富的株高调控基因。中国农业科学院作物科学研究所谷子基因资源团队, 对916 份谷子材料在5 个环境下进行表型鉴定, 将表型数据和重测序数据对接, 在4 个环境下共检测到4 个控制株高的QTL 位点, 分别位于2 号、4 号、6 号和8 号染色体[32]。He 等[33]对谷子育成品种、农家品种和野生种青狗尾草1844 份自然群体进行测序, 并重头组装了110 个染色体级别的基因组, 以此构建了谷子图泛基因组, 结合包括株高在内的多年多点226 组表型数据, 挖掘到10 个与株高相关联的信号。将2 项GWAS 研究株高进行整合分析, 未发现重合的信号, 说明这些GWAS 信号的准确性还有待验证。
除自然群体外, 利用人工构建的群体可以更有目的的发掘双亲中株高控制基因, Feldman 等[34]利用谷子B100 与青狗尾草A10 杂交获得了含217 个自交系的RIL 群体, 利用该群体开发了一张高密度遗传图谱, 结合在人工控制温室的高精度表型鉴定,获得了37 个与株高相关的QTL。由于这是谷子和野生狗尾草之间杂交的RIL 群体, 这些QTL 很可能反映了谷子驯化过程相关的株高控制基因。Mauro-Herrera 和Doust (2016)利用家系数为182 的B100 和A10 的RIL 群体调查了营养生长期、开花期和成熟期谷子的株高表型, 分别获得了7 个、10 个和13 个与株高关联的QTL, 至少在2 个时期可以检测出的有12 个, 其中有7 个位点可在上述2 项研究中可重复检出[35]。经济高效的高通量测序技术辅助开发了更高密度的分子标记, 对RIL 系进行测序并利用bin-map 的方法可以快速得到与表型相关联的最小区间。Zhang[36]利用张谷和雄性不育系A2 杂交构建了包括439 个家系的RIL 群体, 利用重测序数据发掘SNP, 构建了一个2022 个bins 的遗传图谱, 在1号、4 号、5 号、6 号、7 号和9 号染色体, 发掘到6个株高控制位点, 对表型变异的解释率分布为2.94%~44.94%, 其中有的位点也是光周期相关的位点。张谷是个典型的春谷生态型, 而A2 在很大程度上是夏谷类型, 这些QTL 很可能和春夏谷生态型的分化相关。同样是利用A2 和张谷的RIL 群体, Ni等[37]构建了一个含3437 个bins 的遗传学图谱, 在2号和5 号染色体发掘出2 个控制株高的QTL, 其中位于5 号染色体的位点被认为是水稻的绿色革命基因sd1的同源基因, 说明该基因在谷子中同样编码GA20ox 控制株高。Wang 等[38]利用夏谷矮秆品种矮宁黄和春谷品种晋谷21 杂交构建的543 个单株的F2群体, 结合含3129 bins 的遗传图谱, 关联到位于1 号、5 号、8 号和9 号染色体上的7 个与株高相关联的QTL, 解释表型变异率分布为0.39%~30.52%,其中qPH5-2与Zhang 等[36]和Ni 等[37]的株高QTL相重合, 且解释株高变异的30.52%, 贡献率最高。矮宁黄是河北省农林科学院谷子研究所在20 世纪80 年代选育的矮秆品种, 说明水稻绿色革命的矮秆基因sd1在矮宁黄的矮化中起主导作用。He 等[39]利用矮88 和辽谷1 构建的RIL 群体在三亚到公主岭等7 个不同地点调查了333 个家系13 个环境的株高表型, 采用bin-map 的方法定位到26 个与株高关联的区间, 其中13 个QTL 位点可以至少在2 个环境重复检出, 并定位到一个位于qPH1.3内的GA 代谢途径基因, 这可能是控制谷子株高的主效基因。Zhu等[40]利用半矮秆的高度雄性不育系263A 和高秆的创29 杂交, 对F2群体进行高株和矮株的BSA 分析,同时结合父母本节间的表达谱分析, 鉴定出了263A的半矮秆基因很可能是Seita.5G404900, 该基因第6外显子的单碱基缺失, 导致了移码突变, 且该基因是水稻绿色革命基因sd1的同源基因。
谷子株高是一项受多微效基因控制的复杂性状, 尽管株高位点超过了100 个且广泛分布于谷子的 9 条染色体, 但是可以重复检出的位点并不多(表2)。尽管每项研究都挖掘到了谷子株高的QTL的遗传位点, 但由于使用的群体及标记差异, 也只能利用已知信息将部分位点使用统一尺度整合在一起。以Yugu 1 参考基因组物理图谱为准, 在1 号染色体约32 Mb 位置可以重复检测出qph1[36,39], 3 号染色体48.4 Mb 位置可以重复检测出D2[13,34,39], 5号染色体32.6~41.6 Mb 位置可以重复检测出不受光周期影响的qPH5-1/-2[36,38,40], 5 号染色体上的这个位点很可能就是水稻绿色革命基因sd1在谷子上的同源基因。

表2 多环境中检测到的稳定的谷子株高QTLTable 2 Stable QTL for foxtail millet plant height across multi-environments
3 谷子亲本矮88 的株高控制机制解析
矮88 是20 世纪90 年代初期河北省农林科学院谷子研究所利用郑矮2 号为材料采用辐射诱变结合体细胞无性系变异培育的矮秆材料。矮88 的株高较郑矮2 号显著降低, 同时其他株型性状和品质性状也发生了改变。矮88 因紧凑株型受到育种者的重视,很快发展为谷子育种的骨干亲本。到2023 年, 全国利用矮88 及其衍生系为亲本, 培育出审定、鉴定或登记品种83 个, 还有56 个品种完成区试或者正在参加区试, 矮88 成为名副其实的骨干亲本。利用矮88 培育的品种包括冀谷19、中谷2 号、冀谷31、公矮2 号、长生13 等众多在生产上发挥重要作用的大品种, 矮88 后代品种覆盖谷子生产69%的播种面积, 这些品种株高显著降低, 为谷子适应机械化生产做出了贡献。解析矮88 株高控制基因对谷子科研和作物遗传育种研究具有重要的理论和实践意义。
根据表型分析和核心种质的深度重测序基因型分析, 中国农业科学院作物科学研究所刁现民团队挑选出和矮88 没有亲缘关系的高秆品种辽谷1 号与矮88 杂交[41], 历时7 年构建了由333 个家系构成的RIL 群体, 历时4 年在海南三亚、北京、山西长治、河南郑州、吉林公主岭、山西太原、辽宁朝阳等7个地点13 个环境下对群体进行表型鉴定, RIL 群体的株高分布从89.5~149.9 cm, 变异丰富; 表型调查除株高外, 还包括生育期、穗长、穗颈长、节数、叶长、叶宽等, 共获得27 万个数据。对RIL 家系7.88 倍的测序检测到43 万个SNP, 利用这些SNP构建了一个包含3744 bins 的连锁图谱, 图谱的长度为 2101.88 cM, 标记之间的遗传距离为 0.15~37.88 cM, 平均遗传距离为0.56 cM, 是目前报道的谷子上标记最丰富、长度最长的遗传图谱。连锁分析共获得26 个对株高变异解释率2.54%~13.35%不等的QTL, 13 个在2 个或3 个以上环境可重复检测到, 其中包括15 个以前未检测到的QTL。在1 号和9 号染色体, 共发现3 个主效株高QTL 在7~9 个环境下被稳定检测到, 其中qPH1.3效应值2.30%~11.88%,qPH9.2效应值5.07%~10.08%,qPH9.5效应值3.87%~13.35% (图2)。对1182 份谷子核心种质高覆盖测序比较分析发现, 矮 88 在qPH9.2和qPH9.5位点基因型与其他种质均不同, 说明是矮88 特有的矮化基因[41]。这些结果说明矮88 的矮秆表型, 是多个中等或者微效株高控制基因联合起作用的结果, 和安矮3 号、延四直、84133 等单基因控制的矮秆完全不同。利用矮88 为亲本在后代中可以培育不同株高的新品种, 这是矮88 成为骨干亲本的一个原因。
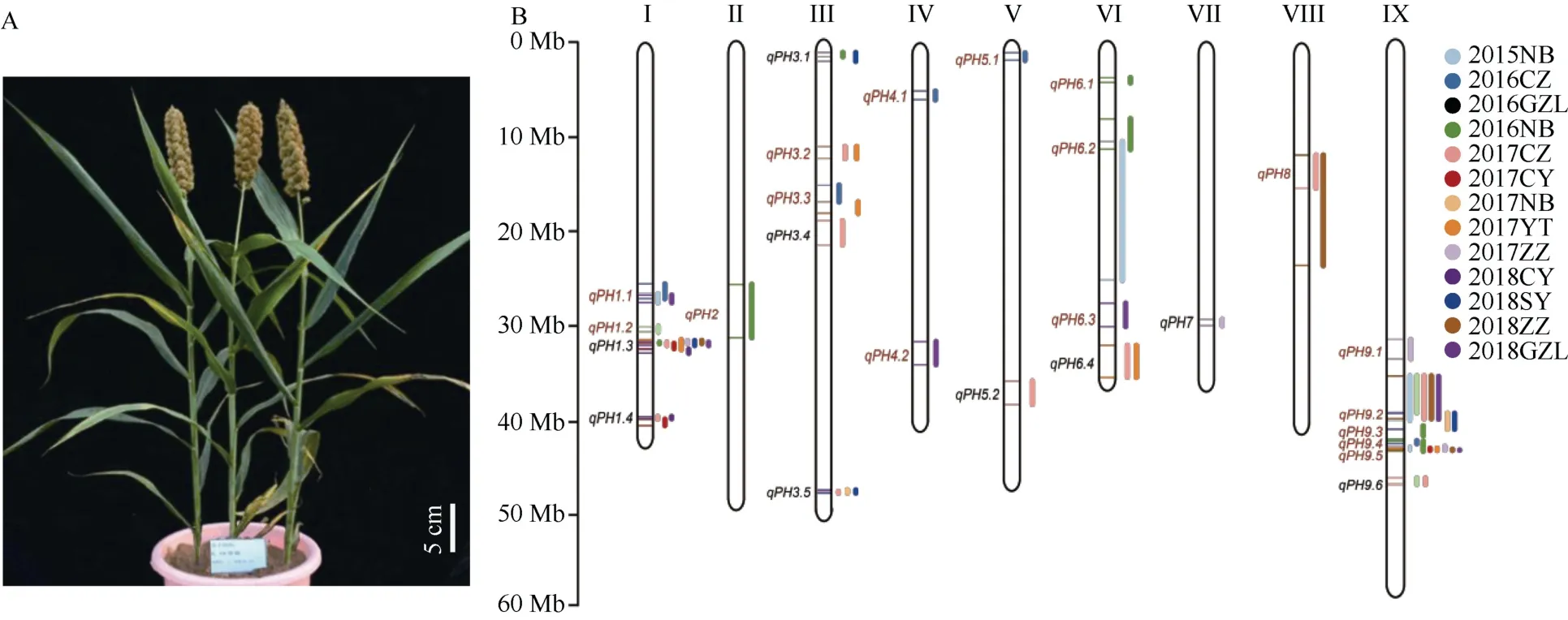
图2 矮88 株高控制基因的遗传解析Fig.2 QTL analysis for foxtail millet plant height using a RIL population derived from Ai 88 and Liaogu 1 (He et al.[39])
对这些QTL 所在区间的基因进行功能预测, 鉴定出6 个和GA 合成或者信号传导相关的基因, 以及15 个编码含F-box 结构域可能和E3 泛素酶关联的基因。特别是在qPH1.3的1.54 Mb 区间, 鉴定出和赤霉素代谢有关的基因, 该基因的活性直接和谷子株高相关, 是谷子育种中选择的株高基因[39]。这些谷子株高控制位点和基因的解析, 对谷子株高精准设计育种至关重要, 也为其他作物的株高育种提供了分子借鉴。
谷穗的长度、穗粗、穗重、穗粒重、一级分枝数(码数)、一级分枝长度(码长)、单码粒数、千粒重等是决定谷子产量的重要因素。利用矮88 和辽谷1 号杂交获得RIL 群体进行QTL 定位, 在13 个环境中共获得了 124 个表型解释率从 0.29%到25.55%的穗部性状相关QTL。其中穗重相关的24个, 穗粒重相关16 个, 穗长相关35 个, 穗粗相关27 个, 码长相关4 个, 码数相关7 个, 码粒数1 个,千粒重相关的10 个[41]。
4 谷子中矮秆基因的育种利用
4.1 矮88 的降秆基因在当代谷子新品种培育中发挥了重要作用
矮秆材料创制和遗传解析的最终目的是培育耐肥水抗倒伏的高产优质品种。虽然谷子育种专家创制了众多谷子矮秆材料, 但能实际在育种中利用的却很少。张家口市坝下农业科学研究所崔文生团队的杜贵等[19]利用矮秆谷子高度雄性不育系构建多父本杂交的动态基因库, 从后代中培育出春谷型的坝矮1 号、坝矮2 号和坝矮3 号等谷子矮秆品种, 其株高分别为77 cm、98 cm 和78 cm, 在高密度栽培下单产达7.5 t hm–2。20 世纪90 年代, 河北省农林科学院谷子研究所培育的矮88 和河南省农业科学院作物研究所培育的豫谷8 号, 是生产上大面积应用的紧凑株型谷子矮秆材料[42-43], 开创了矮秆谷子品种生产大面积应用的先例。姚占廷等[44]利用显性矮秆材料培育出蒙古6 号矮秆品种, 表现抗倒伏、抗病、种植密度由常规品种的45~60 万株 hm–2增加到90~120 万株 hm–2, 产量由4.5~6.0 t hm–2提高到7.5~9.0 t hm–2。
在当代谷子品种培育中, 降低株高的基因以矮88 贡献最大。以矮88 为亲本, 2004 年培育出冀谷19 和公矮2 号2 个中矮秆新品种为标志, 开启了适应机械化生产的谷子中矮秆育种新局面。到2020年采用矮88 和其衍生品种为亲本, 全国谷子主产区育成中矮秆品种115 个。根据对38 个生产中应用的矮88 衍生品种的调查, 华北夏谷区品种株高由135~150 cm 降至100~132 cm, 春谷区品种株高由150~180 cm 降至80~141 cm, 新品种降秆显著。这些矮秆后代品种, 不仅株高显著降低, 而且产量潜力高, 在国家区试中冀谷19、中谷1 号、中谷2号和公矮2 号等7 个品种产量居参试品种的第一位,增产均在10%以上, 说明矮88 是降株高增产量的好亲本; 冀谷40、中谷1 号和中谷2 号突破了生态类型光温反应限制, 在多个生态区表现优异(表3)。冀谷19 在2003—2005 年推广面积居全国夏谷前2位, 并自2007 年连续10 年成为全国夏谷区试的对照品种。冀谷31 在2010—2015 年连续6 年推广面积居全国夏谷第1 位, 年最大推广面积达8.05 万公顷, 是当时全国仅有的2 个推广面积过6.67 万公顷的品种之一。2020 年中谷2 号在山东省推广面积达0.87 万公顷, 居第1 位, 是山东省传统名米龙山小米的主开发品种。
吉林省农业科学院作物研究所利用矮88 为亲本, 培育出公矮2 号, 以及由公矮2 号等衍生的公矮4 号、公矮5 号、公矮6 号等系列矮秆材料, 其中公矮2 号和公矮5 号从20 世纪90 年代中后期开始, 成为了吉林省西部谷子主产区的骨干品种, 满足了机械化管理和收获对矮秆品种的需求。近年来, 用公矮2 号等材料结合拿捕净抗性基因的利用, 培育出公谷85、公谷87 和公谷88 等抗除草剂的矮秆新品种, 进一步提升了谷子轻简化生产的水平, 促进了谷子的大面积产业化发展(图3)。利用矮88 培育的赤谷K3 株高109 cm, 在内蒙古谷子区试中较对照赤谷10 号株高降低46 cm, 但增产4.94%, 提升了该地区中矮秆品种的生产应用和机械化水平。
4.2 中矮秆品种的类型和覆盖率表现出很强的地域特征
东北地区是我国谷子矮秆品种覆盖率最高的地区。2004 年吉林省农业科学院以矮88 为亲本培育出公矮2 号, 开创了东北地区矮秆品种大面积生产利用的先例。2004 年以来, 从公矮3 号到抗拿捕净除草剂的公谷88 等, 已培育出20 多个株高在80~100 cm 之间的矮秆品种在吉林、辽宁和内蒙古东部的谷子生产上利用。根据国家谷子高粱产业技术体系的调研, 矮秆品种在东北地区的覆盖率达63%左右, 是华北、西北和东北3 个谷子主产区中矮秆品种覆盖率最高的地区。包括河南、河北、山东、北京和天津的华北地区则以株高100~120 cm 的中秆谷子品种为主, 如冀谷19、豫谷18、中谷2 号、冀谷31 等。在东北地区生产上起一定作用的矮秆型的吨谷、千斤谷、金谷2 号等, 在华北地区表现中后期长势弱, 容易早衰, 虽然在华北地区也有种植,但面积小。尽管华北地区的主栽品种不是矮秆型,其株高也较原来的老品种显著降低, 老品种株高在135~150 cm, 而新育成品种在100~120 cm。以山西、陕西、宁夏和内蒙古中西部地区为主的我国西北地区, 中矮秆谷子品种的覆盖率最低。这个地区目前以高秆品种晋谷21 为当家品种, 其他高秆品种如晋谷29、汾选3 号、长生7 号和长农35 也有一定面积。利用矮88 为亲本培育的中秆型新品种长生13株高显著降低, 近几年发展迅速, 正在成为这个地区的主栽品种。
公矮2 号、公谷88、金谷2 号和吨谷等矮秆品种在我国东北地区应用较为普遍, 这与东北地区耕地面积大机械化程度高和较为优越的肥水条件相关联; 华北地区的豫谷18、冀谷39、中谷2 号等中秆型品种得到了推广应用, 也基本满足了机械化生产的需求, 保障了以合作社为主要形式的大面积产业化生产的需求。西北地区传统的中高秆品种仍占主导地位, 这可能与西北干旱和丘陵山坡地的生态条件以及谷子产业对品质的要求相关, 西北地区的中矮秆品种在品质上尚达不到传统高秆品种晋谷21、汾选3 号、晋谷29 和8311 的商品品质。西北地区最新培育的长生13 等优质中秆品种, 可以兼顾生态生产条件、抗倒伏和品质的要求, 近年来发展较为迅速。随着我国工业化水平的持续提升, 农业生产的轻简机械化和产业化是不可逆的发展方向, 所有地区谷子新品种培育都必须符合这种趋势。长生13的育成和生产应用, 为西北地区谷子抗倒伏育种提供了矮秆的良好基因资源, 将这个资源和晋谷21 等优质米的性状结合, 必将改变这个地区的品种类型,解决生产中的倒伏和适应机械化生产的问题。
4.3 自然形成的矮秆占有一定的市场地位和份额
在目前生产上应用的矮秆品种中, 除了科研单位培育的矮秆品种外, 一些生产上自然形成的矮秆品种或小型种业公司、合作社选育的矮秆品种也有一定的覆盖率, 如吨谷、千斤谷、旱千谷(旱地千斤)、金谷2 号等, 这些品种的来源系谱多数未知, 他们所具有的矮秆基因也不清楚或不肯定。这些品种来自生产, 具有好的适应性, 加强对这些品种矮源基因的研究, 使他们在谷子矮秆育种和生产上发挥更大的作用。
5 谷子中矮秆基因遗传和育种利用研究展望
5.1 谷子矮秆抗倒研究取得的成果总结
谷子的矮秆种质创新和育种利用以我国为主,国外除印度外, 其他国家基本没有研究报道。我国谷子矮秆遗传和育种研究的成绩可以梳理为以下几个方面: (1) 从20 世纪60 年代开始的谷子矮秆材料种质创新, 积累了数十份各类谷子矮秆资源, 奠定了谷子矮秆种质研究的基础; (2) 从形态上分, 谷子的矮秆材料基本可以分为叶穗下垂的常规型和叶穗直立的直立型; 生物学分析表明, 谷子矮秆材料多数表现早衰严重, 秕谷率高结实差, 限制了矮秆材料的育种应用。造成矮秆早衰的原因与根系弱相关;(3) 对矮秆材料的GA 敏感性分析表明, 除84133 对GA 不敏感外, 其他矮秆材料均对GA 敏感, 说明这些矮秆材料多数是GA 合成途径的突变造成; (4) 遗传分析表明, 多数矮秆材料是隐性单基因控制的,这些矮化基因往往一因多效, 在降低株高的同时也存在着其他不良性状; 已经完成了SiDw1(84133)基因的克隆, 以及SiDw2(T25)、SiDw3(T22)和SiDw4(延四直)的精细定位; (5) 矮88 是中矮秆谷子育种的骨干亲本, 发掘出矮88 株高控制的3 个主效QTL,如果也将这3 个位点的基因命名为矮秆基因, 可以分别命名为qPh1.3(SiDw5)、qPh9.2(SiDw6)和qPh9.5(SiDw7), 共7 个矮秆基因,qPh1.3(SiDw5)等部分矮秆基因和QTL 位点开发了分子标记; (6) 利用创制的矮秆材料, 培育了多个株高显著降低的谷子矮秆或中秆新品种, 很好地解决了谷子倒伏和适应机械化生产的问题; 东北谷子产区以公矮号和公谷号品种为代表, 实现了矮秆品种的大面积覆盖,华北夏谷区实现了中秆品种的全面覆盖, 西北春谷区的长生13 等中秆品种正在迅速发展。
5.2 谷子矮秆和株型研究应注意的问题
首先, 谷子矮秆基础研究薄弱, 限制了矮秆材料的遗传和育种利用。虽然我们积累了众多矮秆材料, 但多数没有进行深入的研究, 只进行了简单的表型观察。加强谷子矮秆材料的遗传和相关基因克隆及其矮化的分子基础解析, 必将为谷子株型育种提供新的遗传基础。已知矮88 的3 个主效矮化位点在育种和生产中发挥了主要作用, 虽然开发了分子标记, 但相关的矮化基因尚未解析; 吨谷、千斤谷和金谷2 号等生产上应用的矮秆品种, 矮化的基因和遗传学基础尚不清楚。加强这些重要矮秆基因的克隆、功能分析和优异单倍型发掘势在必行。
其次, 直立叶片的紧凑株型在水稻、玉米和小麦上都取得了重大成功, 矮化和株型的紧密结合是培育耐水肥超级稻的基础[3-4]。谷子紧凑株型的种质创新和育种利用一直没有受到重视, 更没有显著的进展, 生产上的谷子品种均是披叶松散株型, 株型和群体结构不利于高产和超高产品种的培育, 这可能是谷子产量育种多年来没有大突破的主要原因。科学的将中矮秆和紧凑株型相结合, 才是培育突破性高产谷子品种的关键所在。谷子抗倒伏性受株高和茎秆坚韧度的双重影响[45-50], 在育种中注意矮秆基因和茎秆坚韧度基因的结合, 也需要高度重视。
第三, 矮秆研究如何应对谷子生产的新变化。中矮秆品种的推广很好的适应了机械化收获, 促进了谷子的产业化生产。近年来生产上的一个新变化是干秸收获的面积在快速发展, 东北地区的干秸收获已经占谷子种植面积的40%以上, 华北地区也在快速发展。干秸收获最大的好处是收获后籽粒直接入仓, 或者稍加晾晒烘干就入仓, 减少了籽粒晾晒的用工、场地和设施; 其次, 谷子的联合收割机都是小麦收割机的改装, 谷子籽粒小, 青枝绿叶的活秆收获很容易夹带籽粒, 造成产量损失。干秸收获要求谷子品种成熟后脱水快, 脱水期间谷穗之间不缠绕摩擦落粒, 且植株变干后不倒伏。我国谷子育种家已经开始鉴定和创制后期快脱水的材料, 如何将中矮秆、高坚韧度、紧凑株型和快脱水几个性状进行整合, 是适应产业发展必须面对的问题。
第四, 谷子中矮秆育种应因地而异。株高育种的核心目的是克服倒伏适应高水肥的生产形势。但很多谷子种植在丘陵旱薄地上、山岗地、黄土高原梯田地等, 对于为这些地区培育的品种, 株高不宜过分降低, 因为这些地块以个体和群体结合争取产量, 不同于高水肥地区以群体争取产量。对于华北、东北地区的高水肥地, 中矮秆品种更具有适应性。目前尚缺乏对谷子适宜株高的精准定量研究, 一般认为华北和东北的高水肥地区品种, 适宜株高应在100~130 cm, 而适宜西北的干旱和黄土高原地区,株高以120~150 cm 较为适宜。
5.3 谷子中矮秆基因遗传和育种利用研究展望
虽然谷子矮秆基因的研究和育种利用较水稻、小麦等主要农作物落后, 但谷子可借鉴水稻和小麦的成功经验, 而且谷子的分子遗传和分子生物学近年来快速发展, 作为C4光合作用和禾本科黍亚科的模式作物已被国际学术界接受[53-54], 谷子的高效遗传转化和基因编辑技术体系也已建立, 并在多个谷子功能基因克隆和基因编辑中成功应用, 这为深入研究谷子的矮秆种质创制了平台[24,51-53], 随着矮秆材料遗传基础的解析, 必将提升矮秆材料的利用水平, 也为创制新的矮秆材料提供基础。其次, 谷子高效转化和基因编辑技术体系的建立, 为创制了新的更优良的矮秆材料奠定了基础, 中国农业科学院作物科学研究所在完成谷子高效转化和编辑体系的基础上, 已经建立了基于基因编辑的谷子单倍体诱导技术[54], 这些新技术一方面可直接利用创制新的矮秆基因, 另一方面可以直接对谷子优良高秆品种进行株高关键基因编辑, 直接创制生产可以利用的中矮秆新品种。谷子的泛基因组和图基因组图谱最近的公布, 以及这些基因组变异和多个环境下重要性状的表现型数据的对接, 发掘了大量重要农艺性状的位点和基因, 这为包括标记辅助育种、选择模块育种、全基因组选择育种等方法的实施提供了基础[33]。可以预见, 现代生物技术在谷子中矮秆材料创制和品种培育中必将发挥更大的作用。
猜你喜欢
作物学报(2022年3期)2022-01-22
当代陕西(2021年21期)2022-01-19
麦类作物学报(2021年5期)2021-09-23
农村百事通(2019年17期)2019-10-08
种子(2018年10期)2018-11-14
麦类作物学报(2018年9期)2018-10-09
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
快乐作文·低年级(2016年3期)2016-03-24
