
低氮条件下超级杂交稻苗期根系特征的变化及与其高氮素积累的关系
2024-01-22 06:44崔克辉齐晓丽黄见良彭少兵
作物学报 2024年2期
吴 宇 刘 磊 崔克辉 齐晓丽 黄见良 彭少兵
作物遗传改良全国重点实验室 / 农业农村部长江中游作物生理生态与耕作重点实验室 / 华中农业大学植物科学技术学院, 湖北武汉 430070
水稻(Oryzasativa)是世界上主要的粮食作物之一, 为全球粮食安全做出了巨大贡献。一直以来, 水稻产量增加的同时氮肥投入也在增加[1-2]。然而, 过量施用氮肥往往导致氮肥损失高和利用效率低, 给水稻生产和生态环境的可持续性带来巨大挑战[3-5]。因此, 减少氮肥施用实现水稻绿色生产越来越受到关注[2,4,6-8]。
根系是作物吸收土壤资源的主要器官。研究表明水稻根系形态特征是影响氮肥吸收利用的主要性状[9-12]。陈晨等[13]发现水稻总根长、根体积、根表面积、分枝数与植株的生物量和氮积累量正相关,氮利用效率和产量高的水稻品种通常具有较大根系[14-16], 与氮低效品种相比, 氮高效水稻品种和遗传家系材料具有高的单株根干重、单株不定根根数和单株总根长、最大根长[17-18]。这些结果表明, 提高根干重、总根长、根表面积等根系形态指标有利于促进水稻氮吸收, 这些根形态特征可作为评价水稻氮高效的重要指标。
根解剖结构特征与根氮吸收利用关系紧密,如水稻根系发达的通气组织可促进养分吸收[9],Lynch[19]和Gao 等[20]指出增加根通气组织可以降低根系呼吸消耗, 提高低氮(low nitrogen, LN)下玉米植株氮素利用率。此外, 少的根皮层细胞层数和根系深扎有利于LN 条件下植物生长[21]。研究表明与早期品种相比, 现代小麦品种在生长后期通过维持大的根直径提高氮吸收效率, 根直径与植株氮吸收正相关[22]。类似地, 氮高效玉米品种比氮低效品种具有大的根直径, 这是其适应LN 胁迫的重要根系特征[16]。在LN 下氮高效水稻品种的粗分枝根显著多于氮低效品种, 从而提高LN 下植株养分吸收效率[23]。因此, 根系特征的优化, 尤其是在LN 条件下,有利于植株氮的吸收和利用。
水稻铵转运蛋白(ammonium transporter, AMT)在NH4+吸收中起重要作用[24]。目前在水稻根中发现有10 个铵转运基因表达, 其中AMT1;1、AMT1;2和AMT1;3 属于高亲和铵转运蛋白, 其他属于低亲和铵转运蛋白, 两者在高铵和低铵供应时所起作用大小有所不同[25]。前人研究表明,AMT基因的上调表达显著增加了水稻植株的氮吸收[26-27],OsAMT1;1和OsAMT1;3超表达水稻植株的氮吸收显著增加[25,28-29]。另一方面, 高表达的铵转运基因通过增加叶片光合来促进生长、糖和淀粉合成, 有利于LN条件下干物质积累和产量形成[28]。因此, 铵转运基因的高表达有利于实现水稻高氮素吸收和高产的协同。植物根系所吸收的部分NH4+在根谷氨酰胺合酶(GS)和谷氨酸合酶(GOGAT)作用下被同化为谷氨酸[30], 这2 种酶的活性不仅影响NH4+同化速率,而且与氮吸收和利用密切相关[31]。TaGS2过表达增强了转基因小麦的氮吸收[31],OsGS1;1、OsGS1;2和OsNADH-GOGAT2基因缺失的水稻突变体氮积累和产量显著下降[32-34]。另外, 研究也发现OsAMT1;2和OsGOGAT1同时高表达促进了转基因水稻植株在LN 下氮吸收效率和籽粒产量[35], 氮吸收效率高的水稻品种在LN 条件下根GS 和GOGAT 活性高[36]。因此, 铵同化相关基因的高表达有利于LN 下水稻氮吸收利用。
超级杂交水稻品种比普通水稻品种具有更高的增产潜力[37-38]。与常规水稻品种相比, 超级杂交稻具有总根长大、根表面积大、根系氧化活性高等特征, 从而有利于产量形成和氮吸收利用[9,14]。然而,超级杂交稻高产的实现需要大量肥料和水, 往往导致氮肥过量施用等问题[1-2,4-5]。因此, 在环境友好型水稻生产中实现高产的同时应减少氮肥施用。不同施氮水平影响水稻根系形态、解剖和生理特征, 然而, 超级杂交水稻根系LN 响应特征还不明确。本研究以一个优良超级杂交稻品种和一个常规稻品种为材料, 探究LN 下水稻根系形态和解剖特征的变化及其对氮素吸收的影响, 以期为减氮增效栽培技术研发和新品种选育提供理论依据。
1 材料与方法
1.1 品种选择与氮肥处理
试验在华中农业大学盆栽场室外进行(30.48°N,114.37°E)。供试品种为水稻根系形态和解剖结构差异较大的2 个品种黄华占(Huanghuazhan, HHZ, 常规稻品种, 小根系、根通气组织低)和扬两优6 号(Yangliangyou 6, YLY6, 超级杂交稻, 大根系、根通气组织高)。
试验采用营养液水培法。水稻种子经 10%的H2O2溶液消毒10 min 后浸泡、催芽, 采用水培育秧法。9 d 秧龄时采用国际水稻研究所营养液进行培养[39], 20 d 秧龄时(三叶一心)移栽至塑料容器中(体积为30 L, 600 mm × 360 mm × 150 mm, 桶外壁用锡箔纸包裹隔热)。移栽当天开始氮处理, 设置2 个供氮水平, 即低氮(LN, 0.72 mmol L–1纯氮)和高氮(HN, 2.86 mmol L–1纯氮, 此浓度适宜水稻幼苗正常生长[39]), 使用单一氮源(NH4)2SO4。其余营养元素及浓度为10 mg L–1P (NaH2PO4), 40 mg L–1K (K2SO4),40 mg L–1Ca (CaCl2), 40 mg L–1Mg (MgSO4); 微量营养元素: 2.0 mg L–1Fe (FeCl3)、0.5 mg L–1Mn(MnCl2·4H2O)、0.05 mg L–1Mo (NH4)6Mo7O24·4H2O)、0.2 mg L–1B (H3BO3)、0.01 mg L–1Zn (ZnSO4·7H2O)、0.01 mg L–1Cu (CuSO4·5H2O)和 2.8 mg L–1Si(Na2SiO3·9H2O)。每天用1 mol L–1HCl 或NaOH 溶液调整pH 至5.4~5.5, 每5 d 更换1 次营养液。在相同室外环境下没有移栽水稻植株的营养液中的NH4+含量连续7 d 没有变化, 移栽和没有移栽植株的营养液中没有检测到NO3–。试验设置4 个生物学重复。
1.2 植株干重和氮素含量的测量
选取长势均匀一致的水稻幼苗(6 株)并分为地上部和地下部2 部分, 在80℃烘至恒重并分别称取干重。用MM400 球磨仪(Retsch GmbH, Haan, 德国)将植株样品粉碎, 再置于80℃烘箱烘干至恒重, 混匀后称取4.5~4.8 mg, 利用varioMAX 元素分析仪(Elementar vario MAX CNS/CN, Elementar Trading Co., Ltd, 德国)测定氮含量并计算植株氮素积累量(mg plant–1)。
1.3 根系形态指标测定
氮肥处理后第20 天, 选取长势一致的6 株水稻幼苗, 收集所有根用于测定根形态指标。将根均匀平铺于根盘中并用Epson V700 扫描仪在300 dpi 分辨率下扫描成像, 然后将所有根烘干并称重。用Win RHIZO 2003b (Regent Instruments, 加拿大)进行图像分析, 分别测定根长、根表面积、根体积、根尖数指标。然后, 计算每株的总根长(cm plant–1)、根尖数(No.plant–1)、根表面积(cm2plant–1)、根体积(cm3plant–1)。
1.4 根解剖特征测定
氮肥处理第20 天, 选取植株新长出的不定根(长约8~10 cm), 采用徒手切片法在距根尖2.0~2.5 cm 处切片, 利用尼康Ti 倒置显微镜(Nikon-Ti-SR,日本)在明场条件下成像。用ImageJ 软件(ver.1.39u;NIH, 美国, 网址为http://rsb.info.nih.gov/ij)分析图像, 分别测定通气组织面积(mm2)、皮层横截面积(mm2)、根直径(μm)、中柱直径(μm)、外皮层厚壁细胞厚度(μm), 并计算通气组织比例(根系横切面的通气组织面积占皮层横截面积的比例, %)[40]。每个生物学重复进行6 次技术重复。
1.5 根系铵转运蛋白和谷氨酰胺合成酶、谷氨酸合酶基因表达量的测定
利用TRIzol 试剂(Sigma, 美国)提取根系总RNA,使用微量分光光度计 Nano Drop 2000 (Thermo Scientific, 美国)检测总RNA 浓度和纯度。用DNase I 试剂盒(Invitrogen, 美国)和M-MLV 逆转录酶试剂盒(Invitrogen, 美国)进行反转录。选用SYBR 试剂(Roche Fast Start Universal SYBR Green Master,Roche, 瑞士), 利用ABI Vii A7 Real-time PCR system (Applied Biosystems, 美国)进行实时荧光定量PCR; 具体反应条件: 95℃预变性2 min, 95℃变性10 s, 60℃退火10 s, 72℃延伸20 s, 45 个循环。以水稻Actin基因为内参, 所用引物序列见表1。采用2–ΔΔCT法计算基因相对表达量[41]。

表1 AMT、GS、GOGAT 和Actin 基因扩增引物序列Table 1 The qRT-PCR primers for AMT, GS, GOGAT, and Actin genes
OsAMT基因表达以 HN 处理下 HHZ 的OsAMT1.1表达量作为1,OsGS和OsGOGAT均以HN 处理下HHZ 的OsGS1.1表达量作为1。每个生物学分别进行3 次技术重复。
1.6 数据整理与分析
采用Microsoft Excel 2010、Statistix 9.0 进行数据整理和方差分析, 利用最小显著差异法(LSD)进行平均值差异显著性分析, 2 个品种的4 个生物学重复的平均值用于简单相关性分析。采用SigmaPlot 10.0 进行绘图。
2 结果与分析
2.1 低氮下水稻苗期生物量和氮积累
与HN 相比, LN 处理下HHZ 和YLY6 的根干重分别增加了21%和100%; 然而在2 个氮处理下HHZ地上部分干重没有显著差异, YLY6 在LN 处理下增加了36% (表2)。
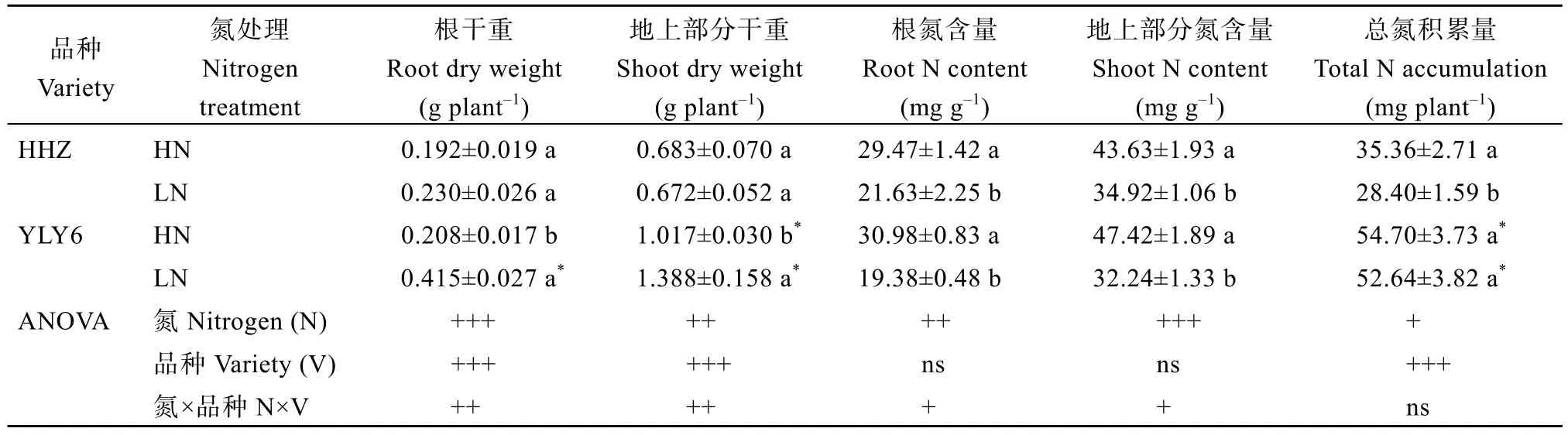
表2 低氮处理对水稻生物量和氮积累的影响Table 2 Effects of low nitrogen application on biomass and nitrogen accumulation in rice
与HN 相比, LN 下HHZ 和YLY6 根系氮浓度分别降低了 27%和 37%, 地上部氮浓度分别降低了20%和32%; LN 处理下HHZ 总氮积累量显著下降,而YLY6 没有显著变化(表2)。在同一氮水平下YLY6的总氮积累均显著高于HHZ (表2)。
2.2 低氮对水稻根系形态的影响
HHZ 总根长、根表面积、根尖数在2 个氮水平间没有显著不同(表3), LN 下YLY6 这3 个表型值分别增加了41%、57%和21%, LN 诱导了2 个品种的根体积显著增加。在LN 下, YLY6 的这4 个形态指标均显著高于HHZ (表3)。
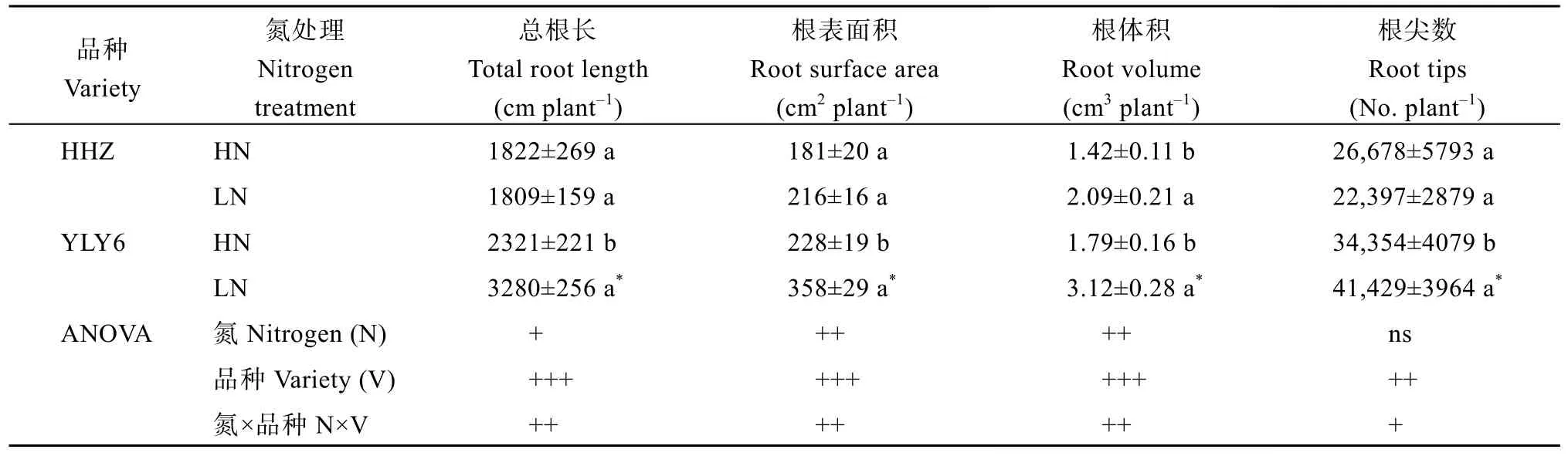
表3 低氮处理对根形态特征的影响Table 3 Effects of low nitrogen application on root morphological characteristics in rice
2.3 低氮对水稻根系解剖结构的影响
与HN 相比, LN 下HHZ 的根直径、根皮层面积和中柱直径分别降低了12.0%、21.9%和11.4% (表4), 而YLY6 分别增加了12.5%、23.1%和2.8%。HHZ和YLY6 的厚壁组织厚度在LN 下分别降低了7.5%和25.2%。LN 下YLY6 的根直径、通气组织比例、根皮层面积和中柱直径均显著高于HHZ (表4)。

表4 低氮处理对根解剖特征的影响Table 4 Effects of low nitrogen application on root anatomical characteristics in rice
2.4 低氮对水稻根系铵转运基因表达量的影响
与HN 处理相比, LN 处理下HHZ 和YLY6 的AMT1;3、AMT2;2表达下降(YLY6 的AMT1;3基因下降不显著), HHZ 和YLY6 的AMT1;3表达量分别下降了73%和33%,AMT2;2表达量分别下降了91.1%和93.3% (图1-C, E)。与HN 处理相比, LN 处理下HHZ 的AMT1;2表达量增加了57%,AMT2;1增加了46%,AMT3;1增加了117%、AMT3;2增加了54%,AMT3;3增加了1420%、AMT4增加了159% (图1-B,D, G~J); YLY6 的AMT1;2表达量增加了 99%,AMT2;1增加了198%,AMT2;3增加了29%,AMT3;1增加了 314%,AMT3;2增加了 390%,AMT3;3(545%)、AMT4增加了105%。2 个氮处理下, YLY6的AMT1;1表达量显著高于HHZ (图1-A); LN 下YLY6 的AMT1;3、AMT2;1、AMT2;3、AMT3;1、AMT3;2表达量显著高于HHZ (图1-C, D, F~H)。HN下HHZ 的AMT2;2表达量显著高于YLY6 (图1-E)。
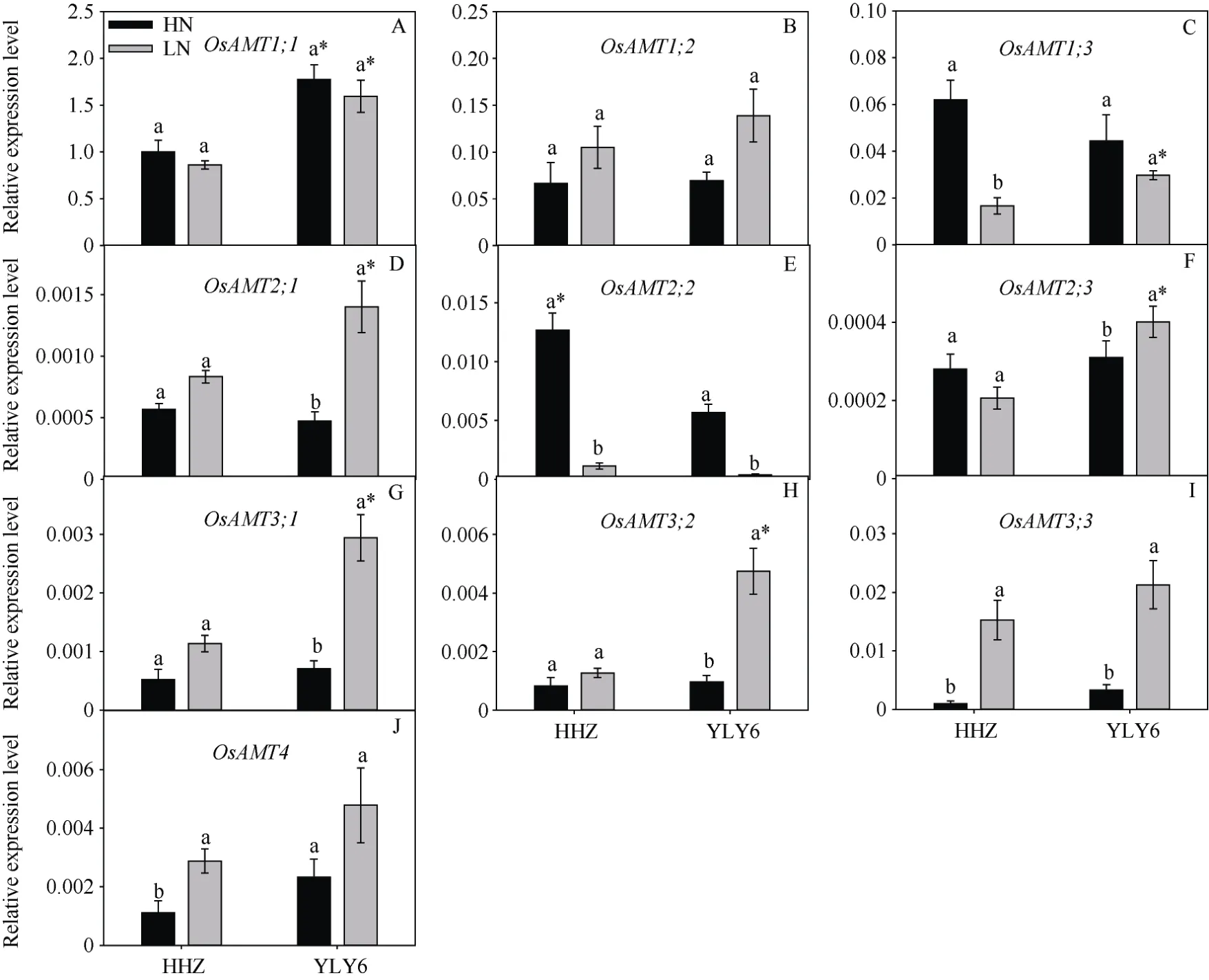
图1 低氮处理对根铵转运基因表达量的影响Fig.1 Effects of low nitrogen application on the relative expression of root ammonium transporter genes in rice
2.5 低氮对水稻根铵同化酶基因表达量的影响
LN 诱导了两品种GS1;1、YLY6 的GS1;3、HHZ的GS2基因表达(图2-A, C, D), 抑制了HHZ 的GS1;2基因表达(图2-B)。在HN 下, HHZ 的GS2表达量显著高于YLY6 (图2-D), 其余铵同化相关基因的表达量在2 个品种间无显著差异(图2); 在LN 下, YLY6的GS1;3表达量显著高于HHZ (图2-C), 而其余铵同化相关基因的表达量在2 个品种间无差异(图2)。
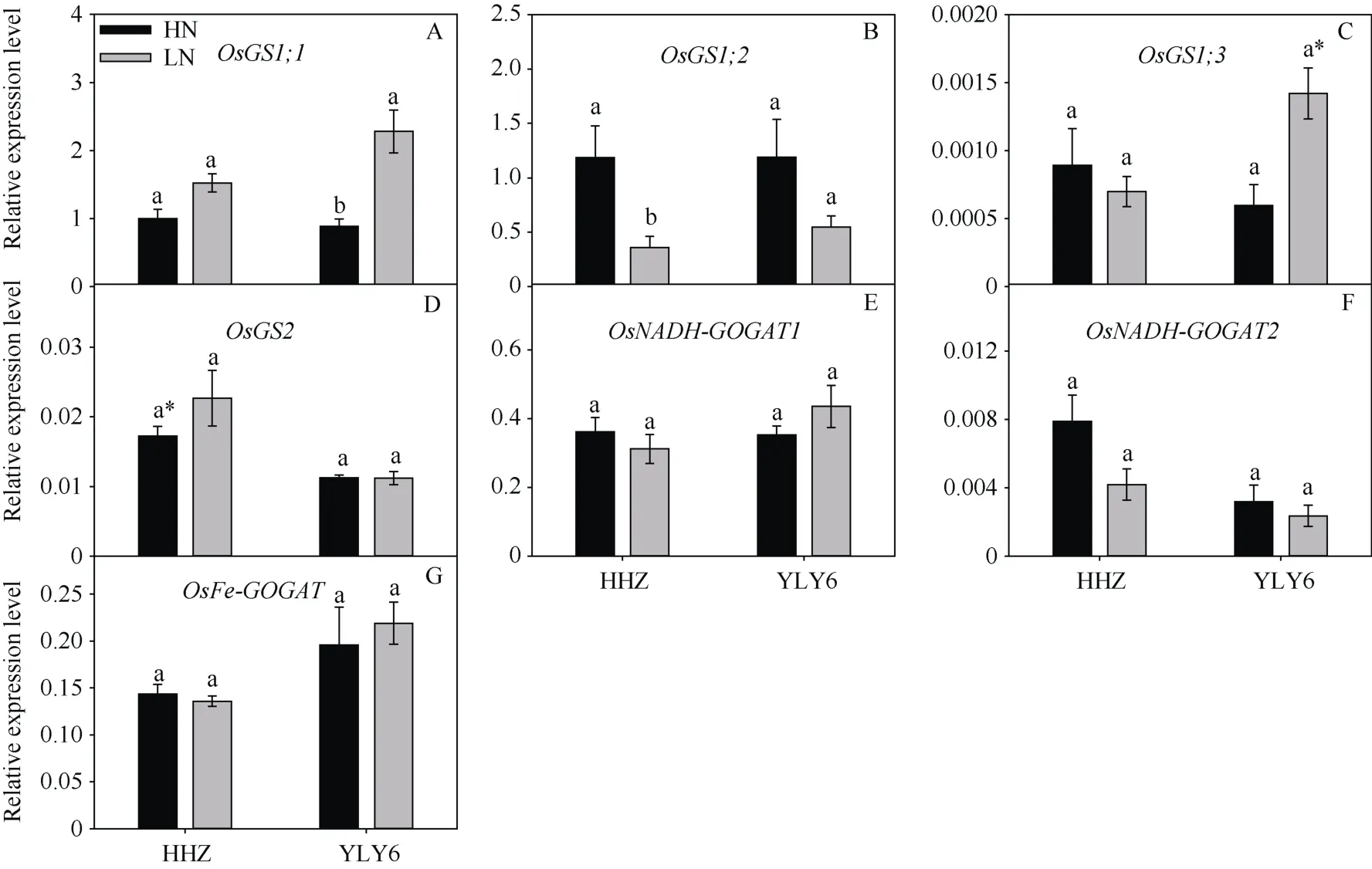
图2 低氮处理对根谷氨酸合酶(GOGAT)和谷氨酰胺合成酶(GS)基因表达量的影响Fig.2 Effects of low nitrogen application on the relative expression of root glutamate synthase (GOGAT) and glutamine synthetase(GS) genes in rice
2.6 根形态解剖特征和相关基因表达与总氮积累的相关关系
HN 处理下总氮积累量分别与根体积(r=0.71,P<0.05)和根厚壁组织厚度(r=0.94,P<0.01)显著正相关(表5), 然而LN条件下总氮积累量分别与所测定的根形态、解剖指标均呈显著正相关(根厚壁组织厚度除外)。
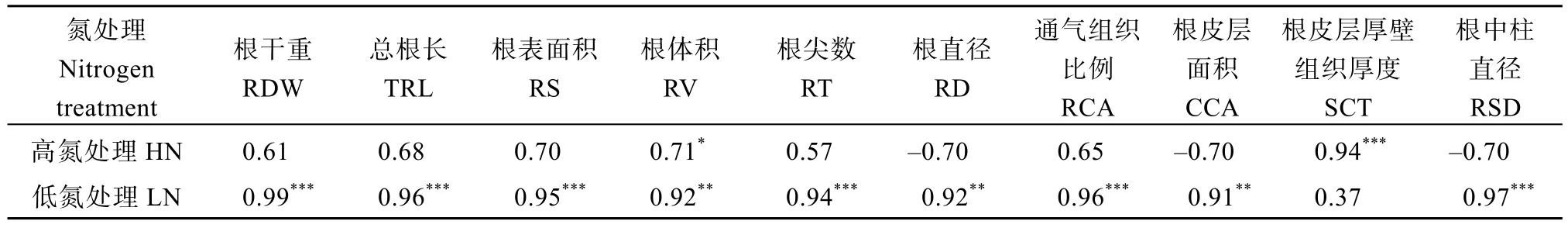
表5 总氮积累与根特征的相关性分析Table 5 Correlation of total nitrogen accumulation with root characteristics
HN 条件下总氮积累量与AMT1;1表达量显著正相关, 而与AMT2;2表达量显著负相关(表6); 在LN条件下总氮积累量分别与AMT1;1、AMT2;3、AMT3;1、AMT3;2表达量显著正相关, 而与AMT2;2表达量显著负相关(表6)。
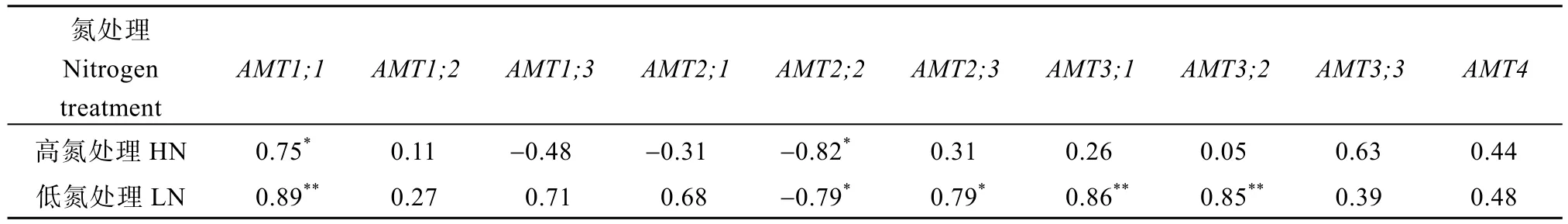
表6 总氮积累与铵转运基因表达量的相关性分析Table 6 Correlation of total nitrogen accumulation with the relative expression level of root ammonium transporter genes
HN 条件下总氮积累量分别与GS2和NADHGOGAT2表达量显著负相关(表7); 然而LN 条件下总氮积累量分别与GS1;1、GS1;3、Fd-GOGAT表达量显著正相关, 而与GS2表达量显著负相关(表7)。

表7 总氮积累与铵同化相关基因表达量的相关性分析Table 7 Correlation of the total nitrogen accumulation with the relative expression level of root nitrogen assimilation related genes
3 讨论
3.1 低氮下水稻根系形态的变化影响氮素吸收
本研究发现2 个氮水平下YLY6 总氮积累显著高于HHZ (HN 下高54.7%, LN 下高85.4%) (表2),并且在LN 下YLY6 总氮积累的降幅也小于HHZ(HHZ 降低了19.7%, YLY6 降低了3.8%), 这结果表明YLY6 具有较高的氮素吸收能力, 尤其是在减氮条件下。值得注意的是, 与HN 相比, LN 处理对HHZ的根干重和地上部干重无影响, 然而显著增加了YLY6 的根干重和地上部干重(表2), 导致LN 下YLY6 的干物质积累高于HHZ。Yuan 等[42]在大田试验中也发现 LN 下 HHZ 干物质积累的降幅高于YLY6。这些结果表明超级杂交稻品种适应LN 胁迫能力可能比常规稻品种强。
与常规稻品种 HHZ 相比, 超级杂交稻品种YLY6 在2 个氮处理下均具有大的根系(表2 和表3)。Chen 等[9]在超级杂交稻中也观察到类似的结果。本研究中LN 下根系形态指标均与总氮积累显著正相关(表5), 这与前人研究结果一致[13]。另外, Liu 等[14]也发现在水稻遗传改良过程中根干重、总根长和根表面积显著增加, 这有利于水稻植株地上部生长和产量提高。Ju 等[43]发现氮高效水稻品种具有高的根生物量和总根长, 杂交稻在各个生育期的根干重、总根长、根表面积、根体积均显著高于常规稻, 这些大的根形态指标是其高氮素积累的重要原因[9,44]。因此, 本研究中YLY6 的大根系可能是其高氮素积累的原因之一。
与HHZ 相比, YLY6 在LN 下根干重的增幅大(YLY6 增加了99.5%, HHZ 增加了19.8%), 同时YLY6 的根长(YLY6 增加了 41.3%, HHZ 降低了0.7%)、根表面积、根体积、根尖数的增幅也显著高于HHZ (表3), 这些结果表明这2 个品种的根形态特征对LN 的响应不同, LN 诱导根特征表型值增加的效应在YLY6 中更明显。Shi 等[36]也发现, 与氮低效水稻品种相比, 在LN 下氮高效水稻品种的总根长、根表面积和根体积增幅更高。前人研究[20]表明与HN 处理相比, LN 可显著提高作物氮吸收效率,主要归因于LN 下总根长、根表面积和根尖数的增加。Ju 等[43]也观察到, 与氮低效水稻品种相比, 氮高效水稻品种在LN 水平下保持高的根生物量和根长, 这些根系特性有利于其在LN 下获得较高的氮素积累和籽粒产量。因此, 本研究中LN 下YLY6 高的氮素积累和低降幅可能与其根系大和根形态特征增幅高有关。
3.2 低氮下水稻根系解剖特征的变化影响氮素吸收
与HHZ 相比, 在LN 下YLY6 具有大的根直径、皮层细胞面积和中柱直径, 同时 LN 显著增加了YLY6 的这3 个性状表型值(表4)。Yang 等[16]观察到LN 下大多数玉米品种的根直径显著降低, 然而与氮低效品种相比, 氮高效品种在LN 下保持大的根直径。Schneider 等[21]发现与根数多且根细的玉米品种相比, LN 下根数少且根粗的玉米品种地上部生长速率和氮积累高。与细根相比, 粗根有利于生育后期小麦氮积累[22]。此外, 与常规水稻品种相比, 超级杂交稻品种具有较高的产量和氮素积累量, 这与其高的根直径有关[14,38]。本研究相关性也表明LN 下根直径、皮层细胞面积和中柱直径与总氮积累呈正相关(表5)。这些结果表明LN 下YLY6 高的根直径、皮层细胞面积和中柱直径可能是其高氮积累的原因之一。
本研究表明LN 显著促进了2 个品种根通气组织的形成(表4), 这与前人研究结果一致[45-46]。LN诱导的根通气组织形成有利于氮积累、干物质积累和籽粒产量形成[9,46-47]。此外, Chen 等[9]发现超级稻品种根通气组织比例高于常规稻品种, 并且这些品种在LN 下也具有高的氮积累。因此, 这些研究表明高的根通气组织是氮高效的根特征之一。值得注意的是, YLY6 在HN 和LN 处理均具有高的根通气组织(表4), LN 下通气组织比例与总氮积累显著正相关(表5)。前人研究表明, 高的根通气组织可降低玉米根呼吸速率和氮浓度, 从而减少了单位根长的代谢成本和构建成本, 更多的碳和氮素分配至地上部,从而有利于植株生长和氮积累[10,46]。因此, LN 条件下YLY6 高的根通气组织可能是其维持高氮素积累量的原因之一。
根细胞壁木质化导致根皮层厚壁组织增厚, 从而抑制根对水分和养分的吸收[48-49]。与HN 相比, LN导致HHZ 和YLY6 根厚壁组织厚度分别降低了7.5%和25.2% (表4)。Kováčik 等[50]发现LN 显著增加了根木质素含量从而不利于离子吸收和积累, Yang 等[51]观察到高NH4+/NO3–处理所致的木质化水平增加导致水稻氮吸收速率降低。另一方面, Ren 等[48]发现降低根木质素含量有利于HN 下水稻水分吸收。这些结果表明, 根皮层厚壁组织加厚和细胞壁木质素沉积可能不利于氮素吸收。因此, 根皮层厚壁组织厚度较大的降幅可能是LN 下YLY6 具有高氮积累的可能原因。
3.3 低氮下水稻根系生理特征的变化影响氮素吸收
水稻铵转运和同化基因表达受到氮水平调节,从而影响氮吸收[25,36,52]。研究表明AMT基因表达上调或过表达显著增强了作物氮吸收[26-29]。在本研究中YLY6 根部分铵转运和同化基因在LN 下的表达量比HHZ 高(图1 和图2)。前人研究发现, 氮吸收效率高的水稻品种在LN 下具有更高的根OsAMT1;1、OsAMT1;3、OsNADH-GOGAT和OsFd-GOGAT表达量[25,36,52]。此外,OsAMT1;1、OsAMT1;2和OsGOGAT1基因的超表达均显著提高了水稻转基因植株氮吸收, 尤其在LN 下[28,35]。本研究观察到LN下铵转运和同化基因的表达(GS2和NADH-GOGAT除外)均与总氮积累正相关(表6 和表7)。这些研究表明在不同氮水平下铵转运和同化基因的表达存在着品种间差异, 这些基因LN 下的高表达有利于氮吸收, LN 下YLY6 根AMT和GS1;3表达水平高可能是其氮积累高的原因之一。
3.4 扬两优6 号根系的低氮响应特征有利于其氮吸收
本研究中LN 处理的施氮量是HN 处理(适宜水稻幼苗生长浓度)的25%, 然而YLY6 和HHZ 的总氮积累在LN 下分别减少了3.7%和19.7%, YLY6 在LN下干物质积累显著增加(表2)。值得一提的是, 本试验每5 天更换1 次营养液, LN 浓度处理下的总氮供应量可能满足了水稻植株生长需求; 同时2 个品种总氮积累都是降低的, 这与其低供氮浓度有关。有研究报道适度LN 处理并不降低水稻植株的生物量,对水稻生长还有显著促进作用[36,43]。Wu 等[12]和Theerawitaya 等[52]发现植株氮积累并没有随着施氮量的增加而线性或成比例地增加, 其可能原因是LN促进了根系生长并优化了根系表型[11-12,53]。此外, 本研究和前人研究都观察到LN 处理诱导了根铵转运和同化相关基因的表达[12,52]。因此, 这些根系性状LN 响应特征有利于氮积累, 适量少施氮肥并不总是降低水稻氮积累和干物质积累。本研究发现LN 下YLY6 氮积累量高和低降幅与其根系形态、解剖特征、铵转运和同化相关基因的LN 响应特征紧密相关。因此, 根系LN 响应特征的评价与优化在氮高效水稻品种选育和减氮绿色生产中应值得关注。
已有研究表明在相同施氮量下超级杂交稻品种YLY6 籽粒产量显著高于常规稻品种HHZ[42,54]; 在氮肥减施50%的大田条件下, 与HHZ 相比YLY6仍然保持较高的总生物量和籽粒产量[42]。另外, Yao等[55]报道YLY6 在不需要高水分投入的同时实现高产。因此, 选育LN 下高氮吸收能力的水稻品种有利于在绿色水稻生产中实现稳产和减氮高效的协同。
4 结论
与HN 处理相比, LN 处理显著降低了常规稻HHZ 的总氮积累, 但对超级杂交稻YLY6 的总氮累积没有显著影响; LN 对HHZ 的根干重、总根长、根表面积和根尖数没有影响, 然而LN 下YLY6 的这些根形态特征显著增加; LN 显著降低了HHZ 的根直径、根皮层面积和中柱直径, 然而LN 下YLY6 的这些根解剖特征显著增加; LN 对HHZ 根AMT2;1、AMT2;3、AMT3;1、AMT3;2和GS1;1基因的表达无显著影响, 然而LN 下YLY6 的这些基因表达显著上调。在LN 条件下超级稻YLY6 的根系形态、解剖和生理特征的响应特征是其总氮积累高于常规稻HHZ 的重要原因。需强调的是, 本研究是在水培条件下进行的, 还需要在田间条件下进一步研究。
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05
四川劳动保障(2021年3期)2021-06-09
动漫界·幼教365(大班)(2018年3期)2018-05-14
现代园艺(2017年13期)2018-01-19
山东农业科学(2017年4期)2017-05-18
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
山东农业科学(2014年2期)2014-03-24
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
