
种植密度对角果期弱光胁迫油菜籽粒油脂积累的影响
2024-01-22 06:44聂晓玉王天尧周元委徐正华波蒯周广生
作物学报 2024年2期
聂晓玉 李 真 王天尧 周元委 徐正华 王 晶 汪 波蒯 婕,* 周广生
1 华中农业大学植物科学技术学院 / 农业农村部长江中游作物生理生态与耕作重点实验室, 湖北武汉 430070; 2 荆州农业科学院, 湖北荆州 434000; 3 宜昌市农业科学研究院, 湖北宜昌 420500
菜籽油是世界上使用量仅次于大豆油的食用性植物油, 也是我国主要的食用植物油之一[1-2]。我国油菜种植区域面积广泛分布于西南及长江流域地区,其中长江流域冬油菜面积占全国90%[3]。据统计,该产区太阳辐射每十年下降1.4%~2.7%[4], 油菜生长期内连续阴雨、光照不足的天气时常发生。遮阴下, 油菜种子数量及重量减少而减产[5]。油菜开花前后遮阴影响生殖器官的发育、碳水化合物累积与转运和籽粒脂质代谢, 从而导致籽粒含油量降低, 蛋白质含量增加[6], 角果期遮阴导致籽粒含油量下降达到44.2%[7-8]。可见, 光照不足已成为限制长江流域产区油菜产量和品质的主要环境因子。
前人围绕弱光下作物的光合作用、物质积累、产量品质做了大量研究。一般认为, 弱光提高了淀粉类种子蛋白质含量, 减少了脂肪含量[9-11]。然而,弱光对油料种子成分的影响更加复杂。有研究表明弱光降低了大豆[12]和蓖麻[13]种子的含油量, 且影响大小随遮阴水平[14]或植株部位不同[15]而变化; 也有研究表明弱光降低了大豆产量, 但不影响种子蛋白质或粗脂肪含量[16]。遮阴改变了田间微环境, 影响了植株的光合进而影响产量形成[17], 而籽粒成分受源库关系的影响[12]。叶片(源)的光合作用产物以可溶性糖的形式输送到果实、种子和根(库)[18]; 可溶性糖通过一系列的生理和生化过程转化为淀粉、蛋白质和脂肪, 并储存在库中, 源强库小有利于粗脂肪的积累。研究表明, 轻度遮阴延迟了油用牡丹‘凤丹’叶片衰老, 增加了光合产量, 通过增加种子重量提高了种子产量, 提高种子发育缓慢生长期和成熟期脂肪酸去饱和酶基因的表达促进不饱和脂肪酸含量累积而改善品质[14,19], 而中度或重度遮光由于光照强度不足, 其光合速率远低于正常光照下的光合速率, 导致光合产量和种子产量降低[14]。水稻中, 籽粒灌浆前期遮阴不仅减少了叶片光合产物形成, 也抑制了源库器官中的蔗糖转运蛋白基因表达, 引起籽粒产量和品质的下降[20]; 棉花中, 遮阴则降低了叶绿素含量、净光合速率、棉花叶片的蔗糖合成酶和蔗糖磷酸合成酶的活性和基因表达也下降, 导致蔗糖含量和蔗糖转化率降低。而且在遮阴的棉叶中观察到较少的淀粉、蔗糖、葡萄糖和果糖含量, 不利于产量和品质的形成[21-22]。在实际生产中, 为了提高弱光胁迫下作物产量、改善品质, 前人围绕施肥等开展了研究, 明确了光照和施肥、种植密度间对作物光合作用存在互作效应[23-25]。遮阴下, 低氮处理下早熟禾植株体内光合产物碳代谢较强, 碳水化合物含量较高; 施氮量增加, 光合产物向氮素代谢转运增多, 碳代谢减弱, 糖含量降低[26]。小麦遮阴下, 适当增加施氮量, 可有效增源扩库, 促进同化物的运输, 导致籽粒淀粉含量显著下降, 蛋白质含量显著增加, 影响籽粒品质[27]。遮阴下, 种植密度过高, 小麦和玉米叶片的叶绿素含量和净光合速率均显著降低[28-29]。
目前, 前人的研究大多集中在弱光胁迫和栽培方式对淀粉类种子生长和产量的影响, 关于种植密度与光照互作对油菜籽粒含油量和油脂累积的研究报道不多见。前期研究表明, 直播油菜密度由传统的低于15 万株 hm–2增加至45 万株 hm–2时, 具有“以密增产、以密补迟、以密省肥、以密控草、以密适机”的效果, 但尚缺乏籽粒品质, 特别是含油量的相关研究[30]。基于此, 本试验模拟光照不足的环境条件, 在油菜角果期进行遮阴处理, 研究种植密度对弱光条件下油菜籽粒含油量的影响, 从籽粒碳、油脂代谢角度, 明确油菜角果期遮阴条件下影响含油量的关键过程, 初步阐明种植密度调控角果期遮阴条件下籽粒含油量、产油量形成机理, 研究结果为光照不足地区油菜耐阴品种的选育和栽培技术的优化提供了理论依据和技术支撑。
1 材料与方法
1.1 试验地概况
试验于2018—2020 年在华中农业大学(湖北省武汉市, 地理位置: 30.52°N, 114.31°E, 海拔23.3 m,亚热带季风性湿润气候)试验田内进行。2018 年土壤养分状况为全氮1.43 g kg–1、碱解氮119.63 mg kg–1、速效磷18.35 mg kg–1、速效钾168.65 mg kg–1;2019 年土壤养分状况为全氮 1.3 g kg–1、碱解氮102.79 mg kg–1、速效磷 20.11 mg kg–1、速效钾188.16 mg kg–1。
1.2 试验设计
选用华油杂62 和沣油520 为材料, 进行大田裂区试验。2 个品种为主区, 3 个种植密度水平[15×104株 hm–2(D1)、30×104株 hm–2(D2)、45×104株 hm–2(D3)]为裂区, 2 个光照强度[自然光照处理(LT-100%)、遮光30%处理(LT-70%)]为裂裂区。油菜群体中50%植株主花序开花时, 在每个小区选取长势一致的植株, 对其主花序当日所开相同部位的花进行挂牌标记, 使用3 层白色60 目遮阴网在10~35 DAF (days after flowering, DAF)遮光, 高度为2.5 m,北侧开放, 方便空气流通; 3 次重复, 共36 个小区。氮、磷、钾、硼肥分别为尿素(含N 46%)、过磷酸钙(含P2O512%)、硫酸钾(含K2O 52%)和硼砂(含B 10.6%)。N 用量为240 kg hm–2, P2O5、K2O 用量均为150 kg hm–2, 硼砂用量11.25 kg hm–2。2018 年、2019年分别在9 月30 日、10 月2 日播种, 分别在次年5月12 日、5 月1 日收获。采用直播方式播种, 行距设定为25 cm。出苗后去窝堆苗, 三叶期至五叶期定苗,小区面积为20 m2(2 m×10 m)。其他同常规管理。
1.3 测定项目与方法
1.3.1 千粒重、单株产量和群体产量 油菜80%左右角果皮呈淡黄色, 籽粒变褐时, 收获并测定小区产量, 同时调查收获株数, 计算单株产量和群体产量; 千粒重采用自动考种分析仪(SC-G 型, 杭州万深检测科技有限公司, 中国杭州)进行测定。
1.3.2 籽粒碳代谢相关生理指标 分别于 25 DAF (遮阴15 d)、35 DAF (遮阴25 d), 在上午09:00—12:00 取各个小区挂牌标记的角果30 个, 室内冰浴(4℃)条件下迅速将角果皮和籽粒分开。籽粒经液氮固定后置-80℃超低温冰箱保存, 用于以下生理生化指标测定。
(1) 叶绿素含量 称取0.2 g 左右籽粒鲜样置于研钵内, 向研钵内滴入95%的乙醇后, 将籽粒研磨至匀浆状并迅速转移至15 mL 离心管中, 向离心管中加入10 mL 95%乙醇密封, 在黑暗条件下, 室温提取18~24 h, 提取完成后摇匀, 在4000 转 min–1的转速下离心5 min, 取上清液, 采用分光光度法测定叶绿素含量[31]。
(2) 蔗糖、果糖、葡萄糖含量 称取0.1 g 左右鲜样, 放入15 mL 离心管中, 以80%的乙醇作为提取液, 在 80℃水浴条件下提取 40 min 后, 于4000×g离心5 min 后取上清液, 以上提取步骤重复2 次, 并以80%的乙醇定容至25 mL。加入0.2 g 活性炭进行脱色, 离心取上清液。具体参考Hendrix(1993)方法测定蔗糖、果糖、葡萄糖含量[32]。
(3) 淀粉含量 80%乙醇提取后的沉淀即可视为淀粉和纤维素的混合物, 因而将淀粉经过糊化反应裂解为单糖的含量换算成淀粉含量。具体过程如下: 乙醇提取残留物在冰浴和避光条件下加入2 mL 浓度为9.6 N 的高氯酸, 15 min 后离心得到上清液, 残留物质加入2 mL 浓度为4.8 N 的高氯酸, 合并上清液即为淀粉裂解为单糖的提取液, 提取液稀释至25 mL。采用蒽酮法测定淀粉含量[33]。
(4) 碳水化合物代谢关键酶活性 采用酶联免疫法测定SuSy、SPS、Rubisco、G6PDH、PPase活性, 试剂盒购于上海晶抗生物工程有限公司(中国,上海)。称取0.10 g 研磨好的籽粒鲜样, 使用1 mL提取液, 进行冰浴匀浆, 离心取上清, 根据制造商提供的说明书进行酶反应。SuSy、SPS、Rubisco、G6PDH、PPase 分别在A480、A480、A700、A450、A700 记录吸收值。
1.3.3 籽粒含油量 油菜籽粒含油量测定参照农业农村部行业标准《油料种籽含量的测定: 残余法》进行。将半径为6 cm 滤纸折成油包, 编号后105℃烘干称重, 记m1; 取约1 g 的籽粒放入油包后包好,105℃烘干称重, 记m2; 将油包放入索氏抽提器抽提8 h, 取出晾干, 105℃烘干称重, 记m3。
1.4 数据处理
利用Microsoft Excel 2016 软件进行数据处理,SPSS 18.0 软件统计分析, 最小显著法(LSD)检验试验数据的差异显著性水平(P<0.05), 采用Origin 9.0软件制图。
2 结果与分析
2.1 千粒重及产量
种植密度增加和角果期遮阴, 均导致两品种千粒重、单株产量降低, 群体产量则随密度增加而增加, 随光照强度减弱而降低。方差分析表明, 品种、遮阴、种植密度对单株产量和产量的影响均达到显著水平, 种植密度和遮阴对单株产量的互作效应达到显著水平。随种植密度增加, 遮阴导致产量降幅逐渐降低。种植密度为D1、D2、D3 时, 遮阴导致华油杂 62 单株产量降幅分别为 31.5%~36.5%、26.5%~30.5%、24.3%~25.6%, 产量降幅分别为31.6%~34.3%、25.0%~28.7%、23.7%~23.8%; 沣油520 的单株产量降幅分别为 31.5%~31.7%、22.0%~22.8%、21.2%~21.5%, 产量降幅分别为27.7%~30.7%、23.8%~23.9%、21.2%~23.1% (表1)。
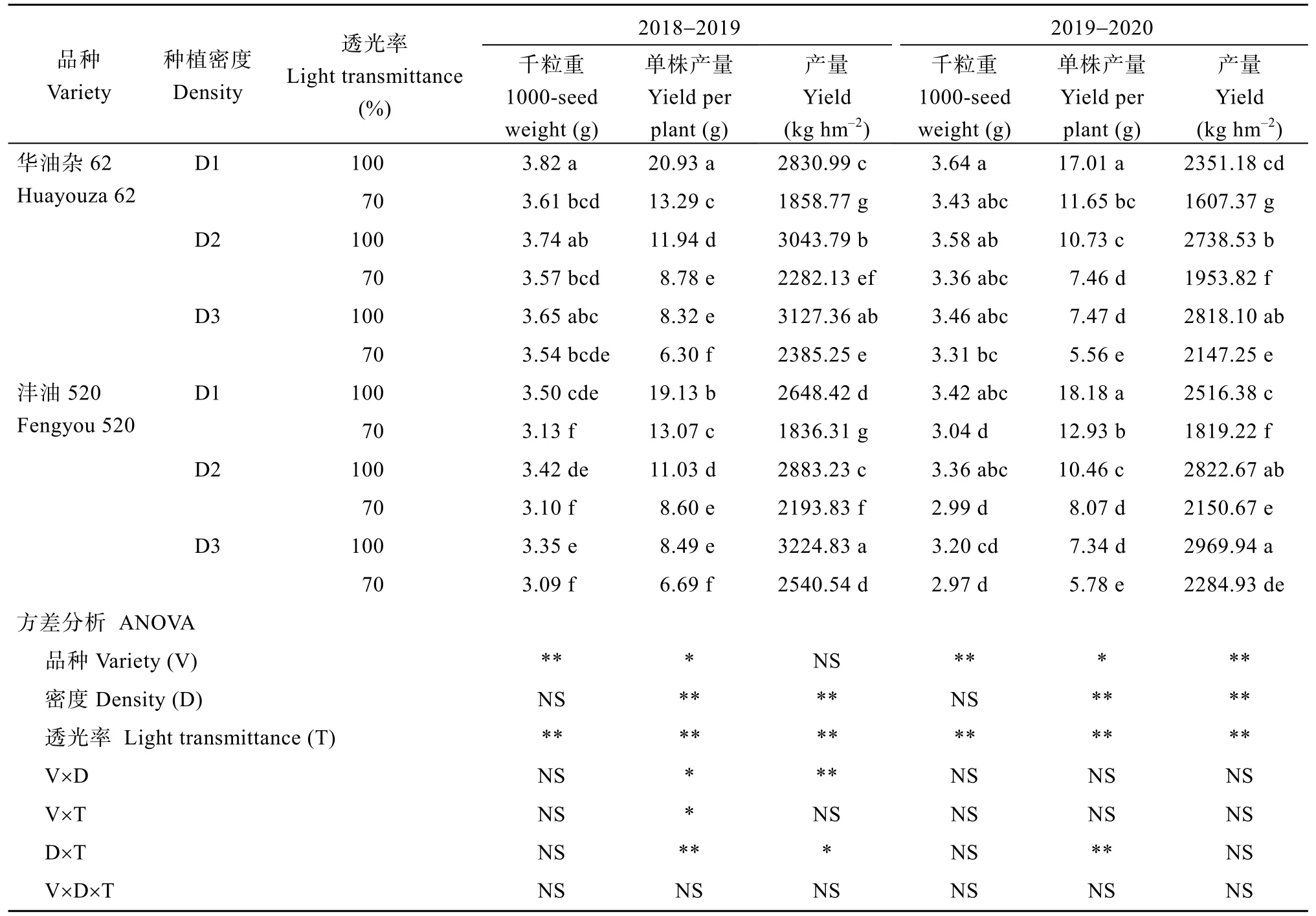
表1 种植密度对遮阴下油菜千粒重及产量的影响Table 1 Effects of planting density and shading on 1000-seed weight and yield of directly seeding rapeseed
2.2 籽粒碳代谢
2.2.1 叶绿素含量 华油杂62 籽粒叶绿素含量高于沣油520。方差分析可得, 种植密度和遮阴对籽叶绿素含量和叶绿素a/b值的影响均达显著水平,花后35 d 时互作效应达显著水平。种植密度增加、角果期遮阴下, 籽粒叶绿素含量和叶绿素a/b值均下降。在2019—2020 年度, 与对照相比, 种植密度为D1、D2、D3 时, 遮阴导致花后35 d 华油杂62籽粒叶绿素含量降幅分别为11.8%~17.1%、9.8%~12.9%、3.3%~4.0%; 沣油520 籽粒叶绿素含量降幅分别为16.1%~19.4%、13.8%~26.9%、8.0%~17.4%(图1)。
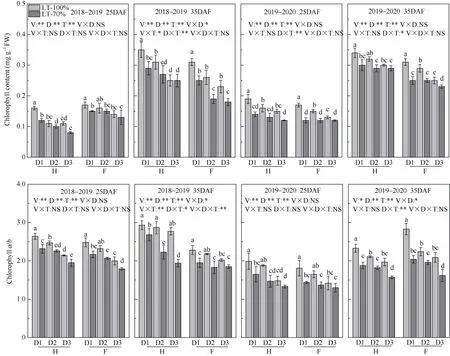
图1 种植密度对角果期遮阴油菜籽粒叶绿素含量和叶绿素a/b 的影响Fig.1 Effect of planting density and pod shading on seed chlorophyll content and chlorophyll a/b ratio of directly seeding rapeseed
2.2.2 碳代谢相关酶活性 华油杂62 籽粒的Rubisco、SPS 和SuSy 活性均高于沣油520。种植密度增加, 花后25、35 d 籽粒的Rubisco 活性降低。方差分析可得, 种植密度和遮阴对籽粒的Rubisco、SPS、SuSy 活性的影响达到显著水平, 且在花后35 d时互作效应达到显著水平。在35DAF 时, 种植密度为15×104、30×104、45×104株 hm-2, 与对照相比, 遮阴导致华油杂62 籽粒的Rubisco 活性下降幅度分别为12.2%~15.2%、9.3%~10.6%、5.6%~8.1%, SPS 活性下降幅度分别为 15.2%~25.1%、9.0%~25.0%、6.5%~19.3%, SuSy 活性下降幅度分别为 12.1%~31.7%、11.2%~29.7%、5.3%~22.8%。而沣油520 籽粒 Rubisco 活性下降幅度分别为 13.8%~17.7%、9.8%~11.4%、4.8%~7.0%, SPS 活性下降幅度分别为17.8%~25.9%、14.8%~28.3%、5.1%~22.7%, SuSy 活性下降幅度分别为16.0%~30.5%、15.3%~25.4%、8.9%~17.6% (图2)。
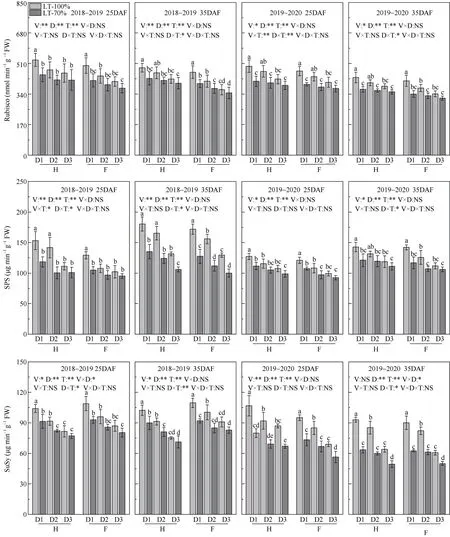
图2 种植密度对角果期遮阴油菜籽粒Rubisco、SPS 和SuSy 活性的影响Fig.2 Effect of planting density and pod shading on seed Rubisco activity of directly seeding rapeseed
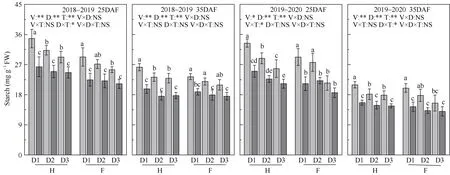
图3 种植密度对角果期遮阴油菜籽粒碳水化合物含量的影响Fig.3 Effect of planting density and pod shading on seed carbohydrates content of directly seeding rapeseed
2.3 籽粒脂质代谢
2.3.1 含油量和产油量 种植密度增加和角果期遮阴, 均导致两品种籽粒含油量下降, 产油量则随密度增加而增加, 随光照强度减弱而降低。方差分析表明, 品种、种植密度和遮阴对2 个时期籽粒含油量、产油量的影响达到显著水平, 花后35 d 种植密度和遮阴对含油量的互作效应达到显著水平。在35DAF时种植密度为15×104、30×104、45×104株 hm–2,遮阴引起的华油杂 62 籽粒含油量降幅分别为4.9%~6.0%、5.6%~7.9%、9.7%~11.8%, 产油量降幅分别为34.7%~35.3%、32.6%~32.8%、27.0%~28.6%;沣油520 籽粒含油量降幅分别为4.1%~5.3%、2.1%~9.1%、6.6%~10.4%, 产油量降幅分别为33.8%~34.5%、29.2%~30.8%、27.9%~30.4% (图4)。

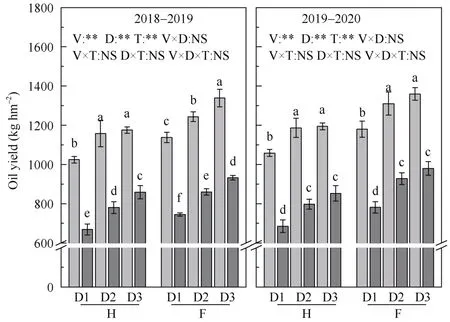
图4 种植密度对角果期遮阴油菜籽粒含油量和油脂产量的影响Fig.4 Effect of planting density and pod shading on seed oil and oil yield of directly seeding rapeseed
3 讨论
弱光胁迫是影响油菜生长和产量的主要非生物胁迫[7], 光合作用下降是弱光下作物生长受限、产量下降的主要原因[25,34]。角果(角果皮和籽粒)是油菜花后重要的光合器官[35], 拟南芥中角果皮的衰老和叶绿素的分解先于种子发生, 籽粒维持光合作用的时间更长[36]。为了使籽粒能够进行光合作用, 靠近角果室内侧具有厚壁细胞层, 具有输送CO2的功能,发育中的种子能够固定这些CO2, 并产生能量用于合成种子的储存物质[37-38], 可见弱光下籽粒碳代谢对稳定其产量和品质具有重要意义。叶绿体是光合作用的重要场所, 叶绿体中的叶绿素是光合作用催化剂, Rubisco 是控制光合作用碳代谢的关键酶, 均受种植密度和光照强度影响[39-40]。本研究结果表明,种植密度增加和遮阴均导致籽粒叶绿素含量、叶绿素a/b比值、Rubisco 活性逐渐降低, 光合产物葡萄糖含量降低; 种植密度增加引起棉花植株间荫蔽程度增加, 植株下层叶片和角果等光合器官的光照辐射减少, 叶绿素合成能力减弱, 光合作用关键酶Rubisco活性降低, 是光合速率下降的重要原因[21]。遮阴后, 油菜角果皮叶绿素b和叶绿素总含量下降,叶绿素a/b值较小, Rubisco活性减少, 净光合速率下降[40], 与本研究结果类似。油菜籽粒在弱光条件下有较低的叶绿素a/b, 使它们能够捕获更多的可用光线[37-38]。优化综合农艺管理措施 (栽培方式、种植密度、施肥管理等)可以调整冠层结构, 增加光截获,减少光损失[41]。光照和种植密度对光合作用存在互作效应。本试验中随密度增加, 遮阴后籽粒的叶绿素含量、叶绿素a/b比值、Rubisco活性下降幅度逐渐减少。而在玉米中, 高密度遮阴后植株的叶绿素含量、叶绿素a/b值下降显著, 光合速率降低[29]。两者结果不同原因可能是玉米中所采用的密度高于所用品种的适宜密度, 而本研究中采用的45万株 hm–2的密度已证实是现有油菜品种密植增产的适宜密度[30],密植改善了角果期冠层结构; 另外, 前期高密度引起的田间荫蔽也提高了油菜角果期耐阴性。
碳水化合物是角果皮和籽粒中的重要物质和能量来源, 并对籽粒的油脂形成和积累有显著影响[42-43]。蔗糖是光合作用的最终产物, 作为主要的转运糖起作用, 植株体内大部分蔗糖被蔗糖合成酶(Sucrose synthase, SuSy)分解为果糖和葡萄糖[44]。果糖和葡萄糖可直接参与糖酵解等代谢途径, 并参与籽粒脂质代谢[45-46]。角果期籽粒虽能通过自身的光合作用合成有机物供籽粒发育, 但前人研究表明十字花科作物中, 籽粒合成的淀粉难以解释观察到的最终油的含量, 需从其他器官中(角果皮、茎秆)转运碳水化合物(如蔗糖和己糖)供给籽粒发育[47]。调控同化物运输到籽粒中的能力越强越利于产量的形成[48], 其中SuSy 在同化物由角果皮向籽粒的转运中具有重要意义[49], 在蔗糖代谢过程中影响库强、具有调控蔗糖的输入和籽粒代谢蔗糖的能力, 从而影响作物产量[50]。前人研究表明, 种植密度增加, 棉花叶片SuSy 和SPS 活性降低, 抑制碳代谢, 蔗糖、可溶性糖含量降低[15]; 而遮阴后, 棉花叶片SuSy 和SPS 活性基因表达和酶活性降低, 导致蔗糖、果糖含量降低[21-22]。本研究中, 种植密度增加或降低光照强度, 25DAF、35DAF 籽粒的SPS、SuSy 活性降低, 降低了蔗糖含量和蔗糖转化率, 导致可溶性糖、蔗糖、果糖含量降低。随密度增加, 遮阴引起的籽粒SPS、SuSy 活性下降幅度减少, 可溶性糖、蔗糖、果糖含量降幅也逐渐减少, 通过增加种植密度, 稳定弱光下油菜籽粒发育、油脂合成的碳源和调节物质, 缓解了弱光胁迫下籽粒千粒重、单株产量和群体产量的下降。
6-磷酸葡萄糖脱氢酶(Glucose-6-phosphate dehydrogenase, G6PDH)控制NADPH 生成量, 是磷酸戊糖途径中的关键酶, 磷脂酸磷酸酯酶(Phosphatidate phosphatase, PPase)是控制三酰甘油合成限制酶, 影响籽粒含油量[51]。弱光下油料类作物籽粒含油量下降[12-13,14-16], 但其机理鲜有报道。种植密度对籽粒品质的影响结论不尽相同。适当增加种植密度, 含油量增加; 同时导致籽粒蛋白质含量降低[52-53]; 但也有研究表明, 种植密度增加, 油菜籽粒含油量减少,蛋白质含量显著增加[54]。密度增加, 油菜籽粒PPase活性上升提高了籽粒含油量[53]。本试验研究表明,同一光照强度下, 种植密度增加, 籽粒的PPase 和G6PDH 活性减少, 抑制籽粒的脂质代谢, 导致含油量降低, 与黄秀芳等[54]结果一致; 且增加密度加剧了弱光对籽粒的脂质代谢的抑制作用, 导致籽粒含油量进一步降低, 但油脂产量因群体产量的增加而增加。方差分析表明, 种植密度和遮阴对籽粒油脂累积关键生理指标在花后35 d 时互作效应达到显著水平。因此遮阴下, 通过高密度种植促进了籽粒碳代谢, 有效缓解了油菜籽粒产量和油脂产量的下降,且花后35 d 是关键调控时期。在油菜生产中, 适当的增加种植密度可显著提高油菜的群体产量从而缓解弱光胁迫下(光照不足地区)产油量的损失。本试验主要从影响籽粒光合作用、碳代谢和油脂代谢的关键物质和关键酶活性方面, 初步阐明种植密度对角果期遮阴条件下籽粒含油量和产油量的影响, 但在角果期, 角果皮是另一重要的光合器官, 其光合特性对籽粒生长发育影响较大, 因此种植密度对遮阴条件下角果皮形态和光合特性的影响有待进一步深入研究。
4 结论
增加种植密度可以缓解油菜角果期遮阴籽粒Rubisco 和SuSy 活性以及果糖和葡萄糖含量的下降,稳定种子碳水化合物的积累, 但加剧了种子G6PDH和PPase 活性的下降, 导致种子含油量进一步下降。然而, 由于群体产量的增加, 产油量的下降随着密度的增加而缓解, 与花后25 d 相比, 花后35 d 是密度和遮光相互作用下影响种子发育的关键时期。
猜你喜欢
农业工程学报(2021年23期)2021-03-11
发明与创新(2019年9期)2019-03-26
安徽科技学院学报(2018年5期)2019-01-18
现代园艺(2017年23期)2018-01-18
种子(2017年12期)2018-01-17
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
动物营养学报(2015年10期)2015-12-01
苏州科技大学学报(工程技术版)(2015年3期)2015-02-28
科学种养(2014年4期)2014-06-09
植物营养与肥料学报(2013年1期)2013-10-26
