
玉米杂交群体产量性状及其特殊配合力全基因组关联分析
2024-01-22 06:44曹言勇
作物学报 2024年2期
马 娟 曹言勇
河南省农业科学院粮食作物研究所, 河南郑州 450002
杂种优势是指F1代杂交种在产量、品质和抗逆等方面优于双亲的一种现象[1]。玉米等作物杂种优势利用在生产和应用上也取得了巨大成功。由于杂交种表型受加性、显性和上位性效应的影响, 因此不能利用双亲表现直接决定杂交种是否优良。1942年Sprague 首先提出一般配合力和特殊配合力的概念, 并用来评价双亲和杂交种表现的优良[2]。特殊配合力(special combining ability, SCA)是指特定亲本组配的杂交后代的平均表现, 用来评价杂交种的实际表现和预测杂种优势[3]。因此, 解析F1杂交种和特殊配合力的遗传机制、挖掘关键控制基因对玉米杂交育种具有重要意义。
全基因组关联分析(genome-wide association study, GWAS)是解析作物复杂数量性状的有效统计方法。近年来研究人员利用GWAS 对F1杂交种和特殊配合力开展了遗传解析工作。Navarro 等[4]利用3552 份F1杂交种对玉米开花性状进行了GWAS 研究, 共鉴定1005 个相关基因。Xiao 等[5]利用42,840个F1杂交种构成的关联群体对单穗重、抽雄期、株高和穗位高进行了遗传解析, 发现上位性互作有助于单穗重和株高杂种优势。利用陕A 群和陕B 群选育的85 个玉米自交系组配的246 份F1杂交种为材料, 李周帅等[3]利用BLINK (Bayesian information and linkage-disequilibrium iteratively nested keyway)
模型对产量和配合力进行了全基因组关联分析, 分别检测到7 个和9 个位点与杂交种产量和SCA 显著关联, 其中4 个为共定位位点。李婷等[6]利用该群体对籽粒性状及其SCA 进行了遗传解析, 分别检测到31 个和5 个显著位点, 其中3 个为籽粒性状和SCA共定位位点。以陕A 群和陕B 群组配的422 份玉米杂交种为自然群体, 李婷等[7]利用加性和显性模型对行粒数、穗长、穗粗等8 个穗部性状进行了关联分析, 分别检测到16 个和3 个显著SNPs (single nucleotide polymorphisms)。Zhang 等[8]利用300 个F1玉米杂交种对株高进行了遗传解析, 共检测到9个显著SNPs 和2 个候选基因。此外, 卢雨晴等[9]以2 份青贮玉米自交系为测验种与92 份自交系组配的184 份测交种为材料, 利用加性模型、显性模型、超显性模型和隐性模型对酸性洗涤纤维进行全基因组关联分析, 定位到35~116 个显著关联位点和64 个候选基因。
尽管以上研究基于F1杂交群体挖掘到一些控制产量、穗部性状和籽粒性状的重要位点和基因, 由于群体背景依赖性限制其不能完全解析F1杂交种和SCA 的遗传机制。因此, 本研究以123 份玉米自交系和8 份测验种按照NCII (North Carolina design II)组配了540 份F1杂交种, 结合8843 个高质量的SNPs标记对该群体单穗粒重、穗部性状、籽粒性状以及其SCA 进行关联分析, 解析F1杂交种产量等性状杂种优势遗传机制, 为选育高产玉米新品种提供参考。
1 材料与方法
1.1 试验材料、田间设计及配合力分析
根据NCII 设计, 利用123 份玉米自交系与8 份测验种(M189、M119、20H1419、S110T、L119A、PH4CV、昌7-2 和农系531)组配了540 个F1杂交种。试验采用随机区组试验设计, 于2021 年种植在河南新乡和周口2 个地点, 1 行区, 2 次重复。新乡小区行长、行距和株距分别是4.00、0.60 和0.22 m, 周口小区行长、行距和株距分别是3.30、0.60 和0.22 m。果穗成熟后, 每小区每个材料选取单穗测量单穗粒重(kernel weight per ear, KWE) (g)、单穗重(ear weight, EW) (g)、行粒数(kernel number per row,KNR)、穗长(ear length, EL) (cm)和穗粗(ear diameter,ED) (cm)。脱粒后, 利用玉米果穗/籽粒考种流水线仪器(国家农业信息化工程技术研究中心)测量百粒重(100-kernel weight, HKW) (g)、粒长(kernel length,KL) (cm)和粒宽(kernel width, KW) (cm)。利用QTL IciMapping v4.2.53[10]计算2 个环境的最佳线性无偏估计值作为F1杂交种表型。利用R 语言lme4 包对两环境联合计算, 获得各性状的SCA 值。
1.2 基因型鉴定及分析
采用CTAB 方法提取123 份自交系和8 份测验种叶片的DNA, 利用玉米5.5K 液相育种芯片(河南省农业科学院粮食作物研究所)进行基因型分型[11]。玉米5.5K 液相育种芯片主要基于探针杂交捕获原理进行基因型分型, 其具体步骤包括: (1) 利用超声打断仪将DNA 片段化并加上测序接头, (2) 将DNA 测序接头与携带生物素标记的RNA 探针进行结合并形成双链复合物, (3) 将链霉亲和素包裹的磁珠和RNA-DNA 双链复合物结合, (4) 清洗获得目标区域DNA, (5) 对目标区域DNA 产物进行PCR 扩增, 构建测序文库, (6) 利用Illumina NovaSeq 6000 进行测序。测序获得的reads 比对参考基因组B73_RefGen_v4 (http://www.gramene.org/), 共筛选到33,971 个SNPs。随后将亲本材料中杂合和缺失的分子标记去掉, 将亲本基因型合并得到F1杂交种的基因型, 并经过最小等位基因频率<0.05 的过滤, 最终获得8843 个高质量SNPs 用于杂交种表型和SCA 全基因组关联分析。123 份自交系和8 份测验种的基因型数据见附表1, 其群体结构和亲缘关系见文献[11]。

表1 不同性状之间的相关系数Table 1 Correlation coefficient among different traits
1.3 全基因组关联分析
基于8843 个SNPs, 本研究利用R 语言GAPIT包的BLINK 模型[12]对540 个F1杂交种的8 个产量性状和其SCA 进行全基因组扫描分析。根据图1, 将模型中主成分个数设置为7。由于杂交种表型受加性和非加性影响, 因此利用加性模型和显性模型分别进行关联分析。GCA 主要受非加性影响, 因此只进行显性模型计算。加性模型的基因编码方式为0、1、2, 显性模型的编码方式为0 和1。以P< 0.05/8843=5.65E-06 为阈值定义显著SNPs。显著SNPs的表型变异解释率利用R 语言GAPIT 包计算。以显著SNPs 上下游各10 kb 为候选区间, 检索Maize GDB 数据库, 同时结合Maize GDB 公共数据库B73穗部、籽粒发育的表达谱数据(表达值≥1)以及注释信息确定候选基因。利用双尾t测验对显著位点不同等位基因型的表型进行差异分析。

图1 主成分分析Fig.1 Principal component analysis
2 结果与分析
2.1 F1 杂交种和特殊配合力表型统计分析
F1杂交种的单穗粒重、单穗重、百粒重和行粒数表型分布、方差分析和遗传力见文献[11]。不同性状之间相关性分析见表1。不管是F1杂交种还是SCA, 百粒重、行粒数、穗长、穗粗、粒长和粒宽均与单穗粒重、单穗重表现出显著正相关关系(r=0.24~0.64)。F1杂交种各性状与其SCA 均具有显著正相关关系(r= 0.61~0.87)。
2.2 杂交种表型和特殊配合力显著关联位点
基于BLINK 加性模型检测到杂交种单穗粒重、单穗重、百粒重、穗长和粒宽显著关联位点各2 个(P< 5.65E-06) (表2)。位点3_229718911 能够解释单穗粒重表型变异的19.62%, 位点2_172799260 解释单穗重变异的81.65%, 百粒重2 个关联位点表型变异解释率为16.62%~19.58%, 穗长2 个关联位点表型变异解释率为11.14%~17.64%, 位于9 号染色体的粒宽显著位点解释27.30%的表型变异, 均为控制杂交种表型的主效位点。
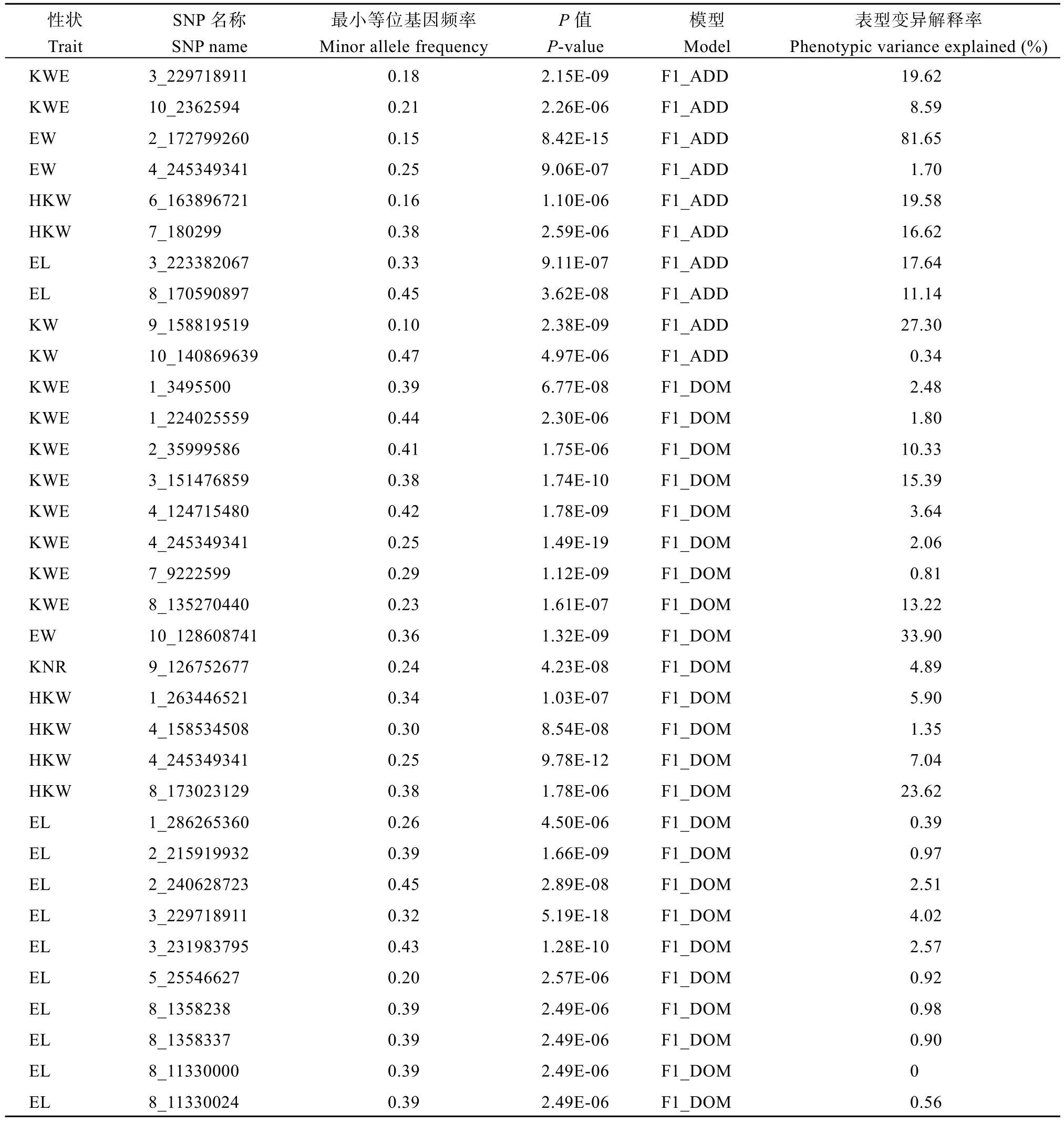
表2 F1 杂交种性状和特殊配合力显著关联SNPsTable 2 Significant SNPs for F1 hybrid traits and special combining ability
利用BLINK 显性模型共检测到31 个F1杂交种表型显著关联SNPs, 8 个与单穗粒重关联, 4 个与百粒重关联, 10 个与穗长关联, 5 个与粒长关联, 6 个与粒宽关联, 单穗重、行粒数和穗粗各检测到1 个显著位点(表2)。与单穗粒重关联的位点2_35999586、3_151476859 和 8_135270440 分别解释 10.33%,15.39%和 13.22%的表型变异, 与单穗重关联位点10_128608741 解释表型变异率为33.90%, 与百粒重关联位点8_173023129 的表型变异解释率为23.62%,说明这5 个显著SNPs 为主效位点。其他显性位点表型变异解释率介于0~7.04%, 均为微效位点。基于BLINK 显性模型检测到8 个显著位点与SCA 关联, 4个与单穗重SCA 关联, 2 个与穗粗SCA 关联, 穗长和粒长SCA 均检测到1 个(表2)。百粒重、穗长、穗粗和粒长各检测到1 个主效位点, 可解释SCA 变异的16.80%~35.86%。
通过比较F1杂交种各性状、SCA 以及不同模型检测的显著SNPs, 发现7 个共定位SNPs (表3)。其中, 3_229718911 为单穗粒重、穗长和粒宽共定位位点。4_245349341 对单穗粒重、单穗重和百粒重表现出一因多效。位点10_128608741 被F1单穗重与其SCA 共定位。与粒宽显著关联的9_158819519 被加性和显性模型同时检测到。

表3 主效和共定位SNPs 挖掘的候选基因Table 3 Candidate genes identified from major-effect and co-detected SNPs
2.3 优异等位基因分析
通过对主效和共定位SNPs 优异等位基因的分析, 本研究发现加性模型均以纯合基因型为有利等位基因, 而且优异纯合基因型几乎均显著或极显著高于杂合基因型, 说明这些位点主要受到加性效应的影响(图2)。对于 F1显性模型, 主效位点8_173023129 和10_128608741 均以杂合基因型为有利基因型, 显著或极显著高于纯合基因型。其中,位点10_128608741 也被单穗重SCA 显性模型检测到,其杂合基因型的SCA 值极显著高于2 种纯合基因型。共定位位点4_124715480 杂合基因型的单穗粒重值介于2 种纯合基因型之间, 但对于粒长而言,该位点杂合基因型为优异等位基因型, 极显著高于最差纯合基因型。对于F1显性模型其余位点, 杂合基因型多表现为最差基因型, 且与纯合基因型也多表现出显著差异, 说明F1杂交种的遗传结构比较复杂, 同时受到加性和显性效应的影响。基于显性模型, 控制穗长、穗粗和粒长SCA 的主效位点均以杂合基因型为优异等位基因, 而且均显著高于纯合基因型, 说明这些位点SCA 以显性效应为主(图2)。但影响单穗粒重SCA 的主效位点4_3412671 以纯合基因型为最优基因型, 杂合基因型表现最差。

图2 主效和共定位SNPs 优异等位基因分析Fig.2 Favourable allele analysis of major-effect and co-detected SNPs缩略词同表1。Abbreviations are the same as those given in Table 1.*: P < 0.05; **: P < 0.01; ***: P < 0.001.
2.4 候选基因的挖掘
为了挖掘对玉米产量等性状及其SCA 具有显著效应的基因, 本研究选择主效和共定位SNPs 进行候选基因挖掘。通过对显著位点位置上下游10 kb范围内检索以及MaizeGDB 数据库B73 籽粒、穗部组织的表达谱分析, 共挖掘到26 个候选基因(表3 和附图1)。通过检索MaizeGDB 和NCBI 数据库, 共获得17 个有注释信息的基因, 包括3 个转录因子(MYBrelated-transcriptionfactor85,NLP-transcription factor9,PHD-transcriptionfactor3), 生长素上调小RNA (smallauxinupRNA11,SAUR11和smallauxin upRNA12,SAUR12), FCS-like 锌指蛋白基因(FCSlikezincfinger16,FLZ16)等。
3 讨论
本研究中F1杂交种产量性状与其SCA 具有较高的正相关关系, 这与前人[3,6]研究结果一致。通过BLINK 模型进行全基因组扫描, 本研究仅检测到1个SNP 同时影响F1单穗重及其SCA 效应。李婷等[6]发现杂交种籽粒表型与SCA 有3 个共定位位点, 占比为9.68%。李周帅等[3]检测到4 个SNPs 为F1产量和其SCA 共定位位点, 分别占F1产量和SCA 显著位点总数的57.14%和44.44%。这些共定位SNPs 表明F1表型与SCA 遗传基础存在相似性。
通过与前人的定位研究结果比较, 本研究中单穗粒重、穗长和粒宽共定位位点3_229718911 位于在Xiao 等[5]利用多个F1杂交群体检测的控制单穗重中亲优势显著QTL 区间内。粒长和粒宽的显著位点10_140869698 和10_140869639 均位于一个控制F1单穗重的显著 QTL 区间内[5]。穗长显著位点 3_223382067 和单穗粒重及其 SCA 显著位点 10_128608741 分别位于Yi 等[13]鉴定的控制单穗重中亲优势显著位点qEWPE3-2 和qEWPE10-3 区间内。与单穗粒重SCA 显著关联的2_2038269 和4_3412671分别位于Li 等[14]定位的F1粒长关联位点(TC-TMMain-qKL2-1、TC-qKL2-1)和百粒重关联位点(TCTM-Main-qHGW4)内。与百粒重显著关联的 7_180299 位于Li 等[14]检测的控制粒长中亲优势位点MPH-qKL7 内。
通过对主效位点和共定位位点进行候选基因挖掘以及MaizeGDB 数据库基因表达谱分析, 共挖掘到26 个候选基因。单穗粒重主效位点8_135270440预测的候选基因Zm00001d010967编码转录因子MYB-related-transcriptionfactor85(MYBR85), 在小麦中TaMYB72是一个调控花期的重要调节因子, 其在长日照条件下过量表达会使水稻花期缩短约12 d[15]。F1单穗重与其SCA 共定位位点10_128608741 对应的候选基因Zm00001d025757编码转录因子NLPtranscriptionfactor9(NLP9)。拟南芥中NLP 转录因子的突变会导致硝酸盐反应和发育过程停滞[16]。单穗重主效位点4_3412671 候选基因Zm00001d048694编码FLZ16, 该基因在雌小穗中偏好性表达[17]。OsFLZ2的共表达会破坏OsMADS51的稳定, 并削弱其下游靶基因Ehd1的转录激活, 从而调控水稻的开花期[18]。穗粗SCA 主效SNP 7_13827332 挖掘的候选基因Zm00001d019039编码转录因子PHD-transcriptionfactor3(PHD3)。大麦基因HvMS1是一个PHD 转录因子, 在花药绒毡层特异性表达, 该基因沉默或过表达会导致大麦雄性不育[19]。对F1单穗重和百粒重具有一因多效性的显著位点1_263446521预测的候选基因Zm00001d033464和Zm00001d 033465分别编码生长素上调小 RNASAUR11和SAUR14在穗原基中高表达(附图1)。水稻中,Os-SAUR33与糖信号通路调节因子OsSnRK1A相互作用,在种子萌发过程中影响糖信号相关基因的表达[20]。拟南芥中,SAUR63会促进下胚轴和雄蕊花丝延长[21]。大豆中, SAURs 在epi-F2:4系中差异表达; 与epi-F2:6相比, epi-F2:4系的产量更高[22]。因此, SAURs 可能是影响F1单穗重和百粒重的重要基因。对候选基因进行挖掘时, 前人将显著SNPs 候选区间设置为1~200 kb[9,23-24]。例如, 卢雨晴等[9]以1 kb 为候选区间检索到64 个控制玉米酸性洗涤纤维的候选基因。吴律等[23]以80 份玉米自交系对玉米穗行数开展GWAS研究, 在5.2 kb 候选区间内检索到4 个候选基因。马雅杰等[24]虽然设置了200 kb 为检索区间, 但最终确定以距离显著SNP 标记最近且具有功能注释的基因为候选基因。为了检索到距离显著SNP 标记最近的候选基因, 本研究利用10 kb 为候选区间并结合基因表达谱, 最终挖掘到26 个候选基因, 同时通过对候选基因家族在其他植物的功能分析, 进一步明确了控制玉米杂交种产量性状最可能的候选基因, 为后续基因功能验证提供指导。
随着分子标记技术和数量遗传学的发展, 研究人员利用双亲、多亲和纯系自然群体挖掘了大量控制玉米产量等重要性状的数量性状位点。但由于玉米商业化生产以单交种为主, 因此基于自交系鉴定的关键位点并不能解释F1杂交种的遗传基础。基于NCII 遗传交配设计的F1杂交群体的亲本材料通常来源于实际育种, 其研究结果可以更好地与育种实践相结合[7]。本研究所用的131 份亲本自交系均是育种材料, 利用BLINK 加性和显性模型鉴定的显著位点可用于该群体的遗传改良。通过对共定位位点优异等位基因的分析, 发现3 个具有一因多效性的位点在多个性状中有利等位基因型表现一致。其中,3_229718911 和4_245349341 为F1不同性状共定位位点, 有利等位基因分别为GG 和TT, 能够累计解释10.80%~23.64%的表型变异(图2)。与单穗重和其SCA 关联的位点10_128608741 的优异等位基因型为GT, 可累计解释37.76%的表型变异(图2)。因此,对这3 个位点开发分子标记可实现多个性状以及F1表型与其SCA 效应的同步改良。NCII 杂交群体通常仅对其双亲进行基因型检测, F1杂交种的基因型则可根据双亲基因型进行推断, 因此对于未组配的杂交组合可利用全基因组选择方法进行预测。基于11,734 个SNPs 标记, Ma 等[11]利用4 种参数模型和一种半参数模型的加性模型和加性-显性模型对本研究群体的单穗粒重、单穗重、百粒重和行粒数进行了预测, 研究发现F1杂交种4 个性状的预测准确性较高, 为0.53~0.75, 可用于有效预测和选择优良杂交组合。因此, 基于NCII 遗传设计组配的F1杂交群体不仅能用于解析玉米产量等重要农艺性状的遗传机制, 还能利用全基因组选择分子育种技术提高优良杂交组合的筛选效率, 加快玉米高产新品种的选育效率。
4 结论
本研究利用BLINK 加性模型、显性模型共检测到38 个和8 个控制F1单穗粒重等8 个性状与其SCA的显著关联SNPs。不同性状和模型之间共定位位点有7 个, 其中1 个为性状与其SCA 共定位。通过对主效和共定位SNPs 候选基因挖掘, 共鉴定到26 个候选基因, 其中转录因子MYBR85、NLP9、PHD3、生长素上调小RNA (SAUR11和SAUR12)、FCS-like锌指蛋白基因FLZ16等可能是控制F1杂交种产量性状与其SCA 的重要候选基因。
附表和附图请见网络版: 1) 本刊网站http://zwxb.chinacrops.org/; 2) 中国知网 http://www.cnki.net/;3) 万方数据 http://c.wanfangdata.com.cn/Periodicalzuowxb.aspx。
猜你喜欢
种子(2023年9期)2023-11-22
西北农业学报(2022年9期)2022-10-08
西南农业学报(2022年1期)2022-03-15
今日农业(2021年14期)2021-11-25
麦类作物学报(2021年3期)2021-05-25
四川蚕业(2020年2期)2020-07-10
北京农学院学报(2020年1期)2020-03-09
科学导报(2018年47期)2018-05-14
中国稻米(2018年1期)2018-03-20
现代农业(2015年5期)2015-02-28
