
Effects of landscape fragmentation of plantation forests on carbon storage in the Loess Plateau, China
2024-02-26 09:11LEIHangyuDUANDantongCHENYiGUOHuifengLIJiangtaoLIXiang
Journal of Arid Land 2024年2期
LEI Hangyu, DUAN Dantong, CHEN Yi, GUO Huifeng, LI Jiangtao, LI Xiang
College of Landscape Architecture and Art, Northwest A&F University, Yangling 712100, China
Abstract: Tree plantation and forest restoration are the major strategies for enhancing terrestrial carbon sequestration and mitigating climate change.The Grain for Green Project in China has positively impacted global carbon sequestration and the trend towards fragmentation of plantation forests.Limited studies have been conducted on changes in plantation biomass and stand structure caused by fragmentation, and the effect of fragmentation on the carbon storage of plantation forests remains unclear.This study evaluated the differences between carbon storage and stand structure in black locust forests in fragmented and continuous landscape in the Ansai District, China and discussed the effects of ecological significance of four landscape indices on carbon storage and tree density.We used structural equation modelling to explore the direct and indirect effects of fragmentation, edge, abiotic factors, and stand structure on above-ground carbon storage.Diameter at breast height (DBH) in fragmented forests was 53.3% thicker,tree density was 40.9% lower, and carbon storage was 49.8% higher than those in continuous forests; for all given DBH>10 cm, the trees in fragmented forests were shorter than those in continuous forests.The patch area had a negative impact on carbon storage, i.e., the higher the degree of fragmentation, the lower the density of the tree; and fragmentation and distance to edge (DTE) directly increased canopy coverage.However, canopy coverage directly decreased carbon storage, and fragmentation directly increased carbon storage and tree density.In non-commercial forests, fragmentation reduces the carbon storage potential of plantation, and the influence of patch area, edge, and patchy connection on plantation should be considered when follow-up trees are planted and for the plantation management.Thus, expanding the area of plantation patches, repairing the edges of complex-shaped patches, enhancing the connectivity of similar patches, and applying nutrients to plantation forests at regular intervals are recommended in fragmented areas of the Loess Plateau.
Keywords: landscape fragmentation; plantation; carbon storage; tree allometry; tree density; structural equation modelling
1 Introduction
In the context of global warming and rising atmospheric CO2content, tree plantation and forest restoration are the leading strategies for enhancing terrestrial carbon sequestration and mitigating climate change (Lewis et al., 2019).To overcome the issue of erosion, China conducted the Grain for Green Project (GFGP) in 1999, which is the largest reforestation operation in the world.After the implementation of the GFGP, the Loess Plateau has undergone fundamental changes in its surface vegetation coverage over the past 20 a, which has had a positive impact on the terrestrial ecological carbon cycle and global carbon sequestration (Liu et al., 2008; Lü et al., 2017).However, forest fragmentation is common on the Loess Plateau (Yang et al., 2018).Planting woodland no longer serves as a single ecological function but is also used as a carbon sequestration method to mitigate future climate change.However, the restoration and growth of plantation forests are affected by landscape fragmentation, which, in turn, affects the carbon storage capacity of forests (Long et al., 2018).Studies have explored the impact of fragmented forest land on stand structure, biomass, and biodiversity, however, the effect of fragmentation on carbon storage in plantation forests remains unclear (Islam et al., 2017; Long et al., 2018; Rolo et al., 2018).
The process of landscape fragmentation is that continuous forests are transformed into small and isolated fragments (Fahrig, 2003).Fragmentation resulted in different negative effects on forests.For example, fragmented forests are highly vulnerable to strong winds, causing branches to break or even die and resulting in a loss of carbon storage (Schwartz et al., 2017).Furthermore,large-scale forest fragmentation threatens species, and species with a high carbon storage potential are replaced by those with a low storage potential (Fahrig, 2003; Chaplin-Kramer et al.,2015).Moreover, trees in fragmented forests are shorter than those with the same diameter in continuous forests (Oliveira et al., 2008; Benítez-Malvido et al., 2017).Fragmentation also intensifies changes in the edge environments of patches (edge effect) (Laurance et al., 2007).Owing to changes in air mixing, solar radiation, and humidity, the microclimate at the edge of a forest is very different from that inside, and is more sensitive to climate change (Schmidt et al.,2017).Forest fragmentation increases the proportion of edge habitats and enhances the impact of microclimatic conditions on carbon pools in fragmented forest patches (Mehmood and Zhang,2001; Laurance et al., 2007; Briant et al., 2010).The edge effect in dry and broad-leaved forests can extend up to 0.8 km inside a forest, and the edge effect on carbon storage exhibits opposite results at different locations (Chaplin-Kramer et al., 2015).Some studies have found that the carbon pool at the edge of temperate forests is higher than that inside the forest, which may be due to an increase in nitrogen deposition and available light and improvement in microclimatic conditions near the edge (Reinmann and Hutyra, 2017).However, an increasing number of studies on temperate forests have shown that the death rate of trees near the edge of fragmented forests generally increases, resulting in a decrease in biomass (Chen et al., 1992; Laurance et al., 1997),and the edge effect may be related to a 10% reduction in carbon density in forests(Chaplin-Kramer et al., 2015).The negative impacts of fragmentation on carbon storage highlights the importance of considering landscape fragmentation when quantifying regional carbon balance (Chen et al., 2004).
The aim of this study was to explore the effects of fragmentation on carbon storage and stand structure of black locust plantation in the Ansai District of Yan'an City, China.We propose the following hypotheses: (1) fragmented forests have lower tree density than continuous forests; and(2) continuous forests have higher above-ground carbon (AGC) storage per hectare than fragmented forests.
2 Materials and methods
2.1 Study area
This study was conducted in the Ansai District, Yan'an City, Shaanxi Province on the Loess Plateau in Northwest China (36°30′45′′-37°19′31′′N, 108°51′44′-109°26′18′′E), which belongs to temperate and semi-arid climate.The average annual precipitation is 505.3 mm, which concentrated in July-September.The annual average temperature ranges from 7.7°C to 10.6°C,with the lowest temperature of -24.0°C in winter and the highest temperature of 37.0°C in summer.The annual evaporation is more than 13 mm, and the average accumulated temperature is 3160.2°C.The average frost-free period is 157 d, and the total annual sunshine is 2415 h.The soil in the study area is mainly loess, and the soil texture is powdery or silty, which has good water permeability.The government of China started returning farmland to forests in this area in 1999.The black locust (Robinia pseudoacaciaL.),Pinus tabulaeformisCarrière, andPlatycladus orientalis(L.) Franco ex Hu & W.C.Cheng are the main plantation species (Yang et al., 2022).
2.2 Experimental design
Plantation forests with similar growth ages, ecological environment, and human disturbance were selected for this study.Based on these criteria, we excluded small patches (<2 hm2) from the selection of fragmented forests because they were more easily disturbed than large patches.This study also reduced the choice of extra-large patches (>15 hm2) to prevent fragmented patches,similar to continuous patches.In this study, the choice of fragmented patches was concentrated at medium sizes (3-15 hm2).
The study was conducted in July 2022, in which total 44 samples (22 for fragmented and 22 for contiguous forests) with an area of 10 m×10 m were set up in fragmented and continuous plantation forests (Fig.1).All samples were placed at least 50 m from the nearest forest edge to avoid human disturbance, and obvious canopy gaps and areas with signs of human disturbance were avoided.There was a distance of at least 300 m between samples to reduce the interference(Fuller and Enquist, 2012).
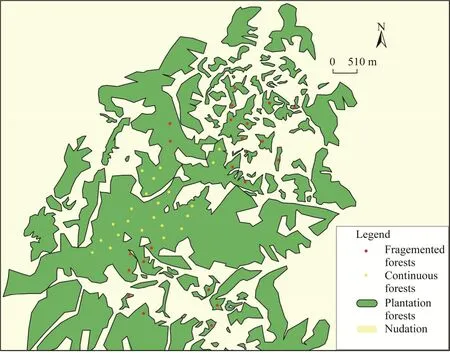
Fig.1 Sampling locations in fragmented and continuous forests of the study area
2.3 Data processing
Since the implementation of the GFGP in 2009, replanting measures have been undertaken.This study measured and recorded the trees with a diameter at breast height (DBH) >9.0 cm and calculated the number of trees and DBH using a tape measure to eliminate the artificial influence on tree density.Height from the ground to the top of the canopy was measured using a laser rangefinder, and if the tree had multiple branches/stems at 1.3 m height, each branch diameter >9.0 cm was recorded, and the DBH of these multi-stemmed branches was estimated.The presence or absence of canopy coverage was recorded every 1.0 m along two adjacent sides of each 10 m×10 m plot, with presence recorded as 1 and absence recorded as 0.The percentage of presence was used as the canopy coverage (Surendra et al., 2021).The total number of trees recorded for each sample plot is considered as the forest tree density for that plot (trees/0.01 hm2)because of the consistent planting densities at the time of fallow.The size of the tree densities surveyed in this study can be used as a measure of how well trees are surviving in both landscape types (Davies-Colley et al., 2000).The DBH and height of trees in each sample plot were averaged as the DBH and height of the sample plot, and only trees with a DBH>9.0 cm were recorded.
2.4 AGC stocks
The biomass of each tree was calculated using the black locust growth model fitted by Li and Liu(2014) in the Ansai District (Table 1).The sum of these values was multiplied by the carbon fraction (0.5) to obtain the total AGC for each sample plot, which was scaled to give an estimate per hectare.
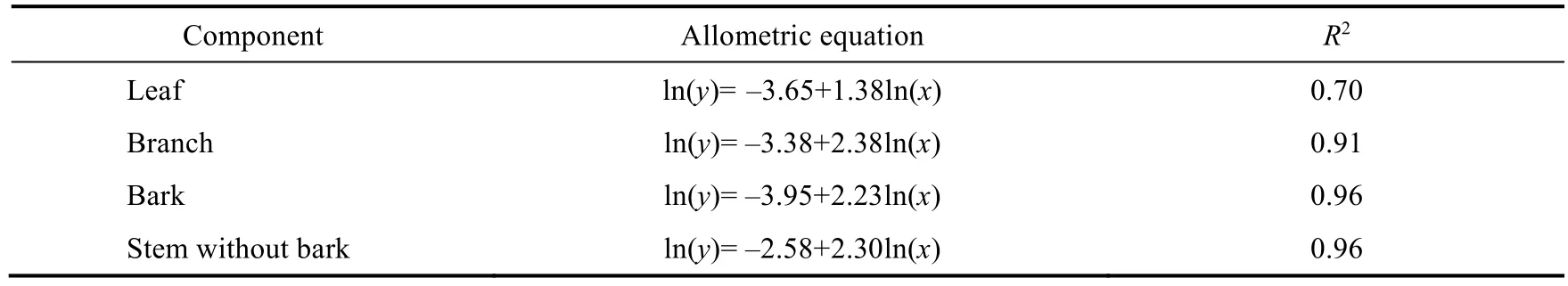
Table 1 Allometric equations for estimating each component biomass of black locusts in the Loess Plateau
2.5 Land cover and fragmentation
Visual interpretation of height definition satellite maps using ArcGIS v.10.6 software (Fig.1) was used to distinguish the forest plots from nudation.A circular map with a radius of 300 m was cut at the centre of each sample, and the landscape index was calculated using the Fragstats v.4.2 software.Three landscape indices were calculated to represent the three key axes of fragmentation at the patch scale, i.e., size, edge, and isolation (Fig.2; Schwartz et al., 2017).The patch area (AREA, hm2) represents patch size.The edge using shape index (SHAPE) was calculated as follows:
wherepis the circumference of the patch (m); andais the surface area of the patch (hm2).SHAPE increases with an increase in patch circumference, and if the patch is perfectly conformal,it is equal to 1.We quantified isolation using proximity index (PROX), which considers the area and distance of patches of the same type within a specific radius around the study patch and increases from zero, with an upper limit determined by the search radius.For a given patchi, the PROX is calculated as follows:
whereaijis the area (m2) of focal patchiwithin a specified radius of the same type of patch(j=1, …,n);hijis the distance between patchesiandj(m); andnis the number of patches of the same type with focus patches around a radius of 300 m.This equation indicates that larger and closer patches of the same type reduce the isolation between patches more than smaller or more distant patches.Because the minimum distance between the samples was 300 m, the neighbourhood radius was set to 300 m.At the class scale, patch density (PD; number of patches/100 hm2) was used to describe the degree of fragmentation at a radius of 300 m (Fig.2),which reflects the intensity of human or natural disturbance to the landscape; the greater the degree of fragmentation, the lower the landscape security.PD is calculated as:
where PD is the patch density (number of patches/100 hm2) of type I;Niis the number of patches of type I; andAis the total area (hm2).

Fig.2 Conceptual diagram illustrating axes of fragmentation and variables associated with fragmentation included in analyses.Green squares represent forest pixels, and adjacent pixels represent a patch.The red outline indicates a focal patch for isolation measures.
2.6 Topographic wetness index (TWI)
The impact of abiotic factors on plantation forests was characterised using TWI, which has proven to be a reliable predictor of tree growth, as demonstrated by studies on species such asQuercusspp.(Petroselli et al., 2013).Its significance is highlighted in the plantation forests of the Loess Plateau, where variable topography influences forest distribution, and limited climatic variations across the study landscape make it particularly relevant (Mucina and Geldenhuys,2006).TWI calculations were conducted using a 30 m resolution digital elevation model with ArcGIS v.10.6 software.This index quantifies an area's water accumulation capacity, with lower values indicating reduced capacity in steep areas.Higher values are typically associated with flat areas prone to water accumulation.TWI was calculated for each pixel using the following formula:
whereAsis the drainage area (m2); andβis the slope (rad).
2.7 Distance to edge (DTE)
Edge effects have been shown to have a significant impact on forest lands (Miranda et al., 2022;Willmer et al., 2022), with large changes in light temperature, moisture, and air in the forest as the DTE becomes progressively smaller (Pöpperl and Seidl, 2021; Zhang et al., 2021).We characterised the distance from each sample point to the DTE (Paulson et al., 2021) and placed it as an independent variable in the structural equation model (SEM) to quantify the edge effect.DTE was calculated using the nearest neighbour distance tool in the ArcGIS software.
2.8 SEM
We used SEM to test the direct and indirect effects of the landscape metrics, TWI, DTE, and forest structure on AGC (Fig.3).Our conceptual model was based on previous studies reporting the direct and indirect effects of landscape and abiotic metrics on forest carbon pools (Andivia et al., 2016).The standardised data were fitted to a structural equation model using the maximum-likelihood approach (Table S1).Using Stata's own goodness-of-fit statistics, the chi-square (chi-square goodness of fit test) was selected to assess the model's goodness of fit, andP<0.05 represents a good model fit.The fit of each equation was tested using Stata's command.We ran the principal component analysis (PCA) and retained the first component as a surrogate for the landscape metrics to reduce the dimensionality of the landscape metrics and avoid collinearity.The first component explained 50.3% of the variance and was positively related to AREA (loading is 0.89) and negatively related to the total amounts of SHAPE, PROX, and PD(loadings are -0.62, -0.11, and -0.92, respectively).We reported the standardised path coefficient of each significant link to allow comparisons across variables of varying units.Standardised path coefficients can be interpreted as the expected change in the response variable (e.g., AGC) as a function of the change in predictor (e.g., fragmentation) in standard deviation units.We also report the conditionalR2values of the response variables (i.e., the variance explained by fixed and random effects).All statistical analyses were performed using Stata v.17.0.
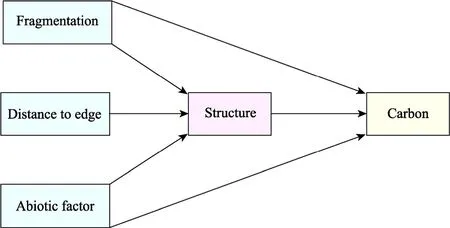
Fig.3 Conceptual model describing how fragmentation and environmental attributes relate to forest structure and how all of these attributes relate to carbon storage.To test the conceptual model, we used patch area, edge,and proximity index with patch density as proxies for forest fragmentation, topographic moisture index (abiotic factor), distance to edge, canopy coverage, and tree density (structure), and above-ground carbon storage(carbon).
2.9 Data processing and analysis
Thet-test was used to test the significant difference between fragmented and continuous grouped data (P<0.050), using the unpaired and assuming data as Gaussian-distributed options for the fragmented and continuous groups.AREA, SHAPE, and PROX did not conform to a Gaussian distribution.All three landscape indices were logarithmically treated, and a linear regression model was used to fit the four fragmentation indices to the unit sample carbon storage.To explain the anisotropic relationship between black locusts in fragmented and continuous forests, we fitted the height and DBH data using linear regression for all trees in the two groups.Carbon storage calculations were performed using Microsoft Excel, andt-tests and linear regression analyses were performed using GraphPad Prism v.9.4.1.
3 Results
In this study, 455 black locust plantations were surveyed in 44 sample plots in two types of forests and use independent samplet-test to test the significant difference of 6 groups of data collected in 44 quadrats of fragmented and continuous forests.
3.1 Black locust forest structure and carbon storage
Regarding the average DBH of the sample plot, the black locusts in fragmented forests was 53.3%thicker than that in continuous forests (P<0.010; Fig.4a; Table 2).However, height did not differ(P=0.140; Fig.4b; Table 2).Carbon storage in fragmented forests was 49.8% higher (P<0.010;Fig.4c; Table 2), but tree density was 40.9% lower than those in continuous forests (P<0.010; Fig.4d; Table 2), and canopy coverage in continuous forests was 20.0% higher than that in fragmented forests (P<0.010; Fig.4e; Table 2).
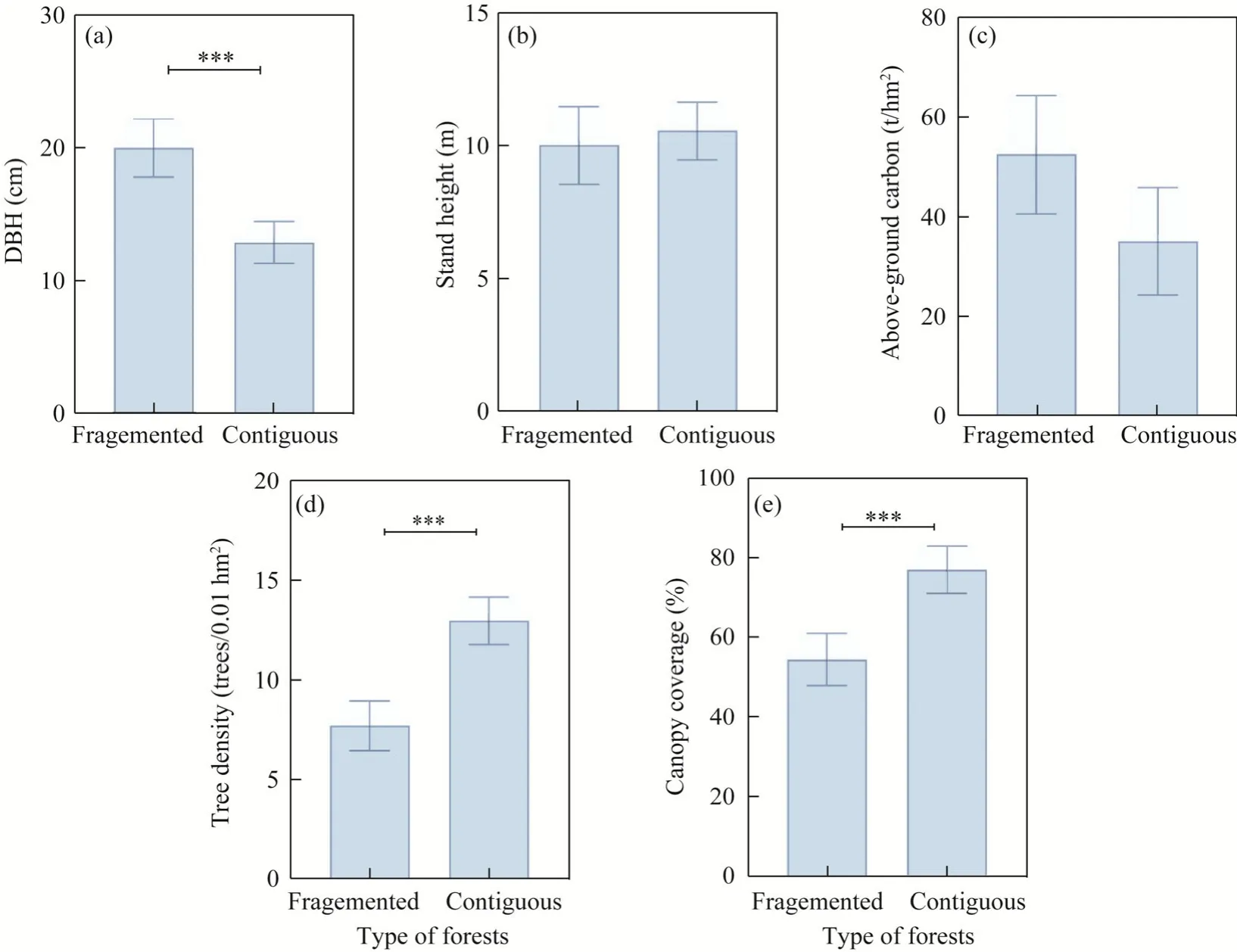
Fig.4 Forest structure under fragmented and continuous forests.(a), DBH (diameter at breast height); (b), stand height; (c), above-ground carbon; (d), tree density; (e), canopy coverage.Bars are standard errors, and vertical line represents 95% confidence interval.***, P<0.001 level.
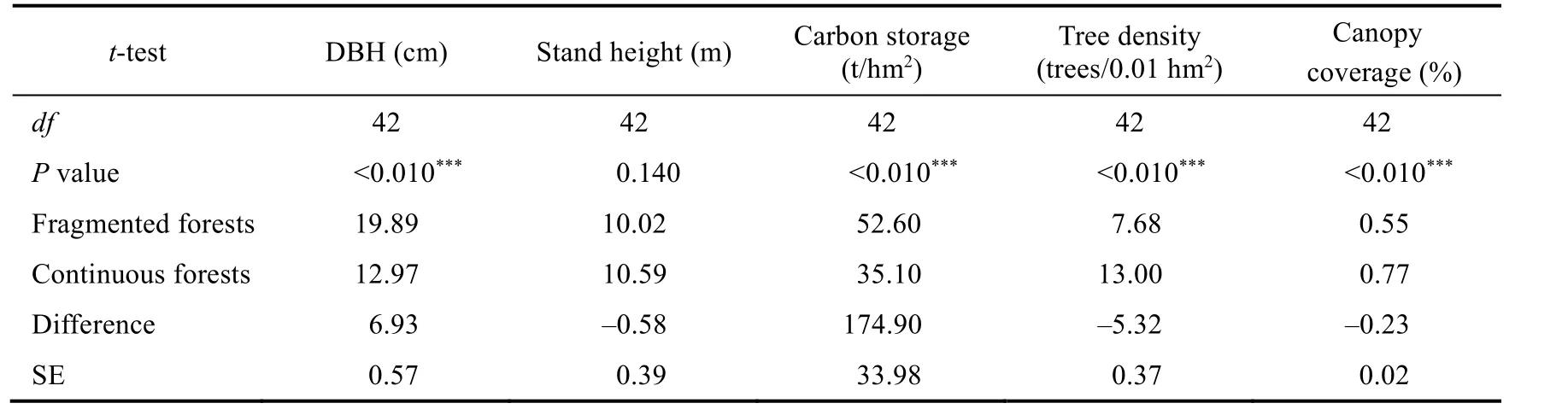
Table 2 t-tests for DBH, stand height, above-ground carbon storage, tree density, and canopy coverage for two types of forests
3.2 Relationship between tree height and diameter
When DBH of black locust was >10 cm, the trees in continuous forests were taller than those in fragmented forests.When the DBH was <10 cm, the trees in fragmented forests were taller than those in continuous forests.With an increase in DBH, the height of trees in fragmented forests did not change significantly and remained at approximately 10 m (Fig.5).However, the height of trees in continuous forests increased significantly (Fig.5).
3.3 Relationships between four types of fragmentation indices and carbon storage
AREA exerted a significant effect on carbon storage.With an increase in AREA, the carbon storage decreased (Fig.6a).Edge, isolation, and PD had no significant effect on carbon storage(Fig.6b-d).

Fig.5 Relationship between stand height and DBH (diameter at breast height) under fragmented and continuous forests.Shaded area represents the 95% confidence interval of the regression.
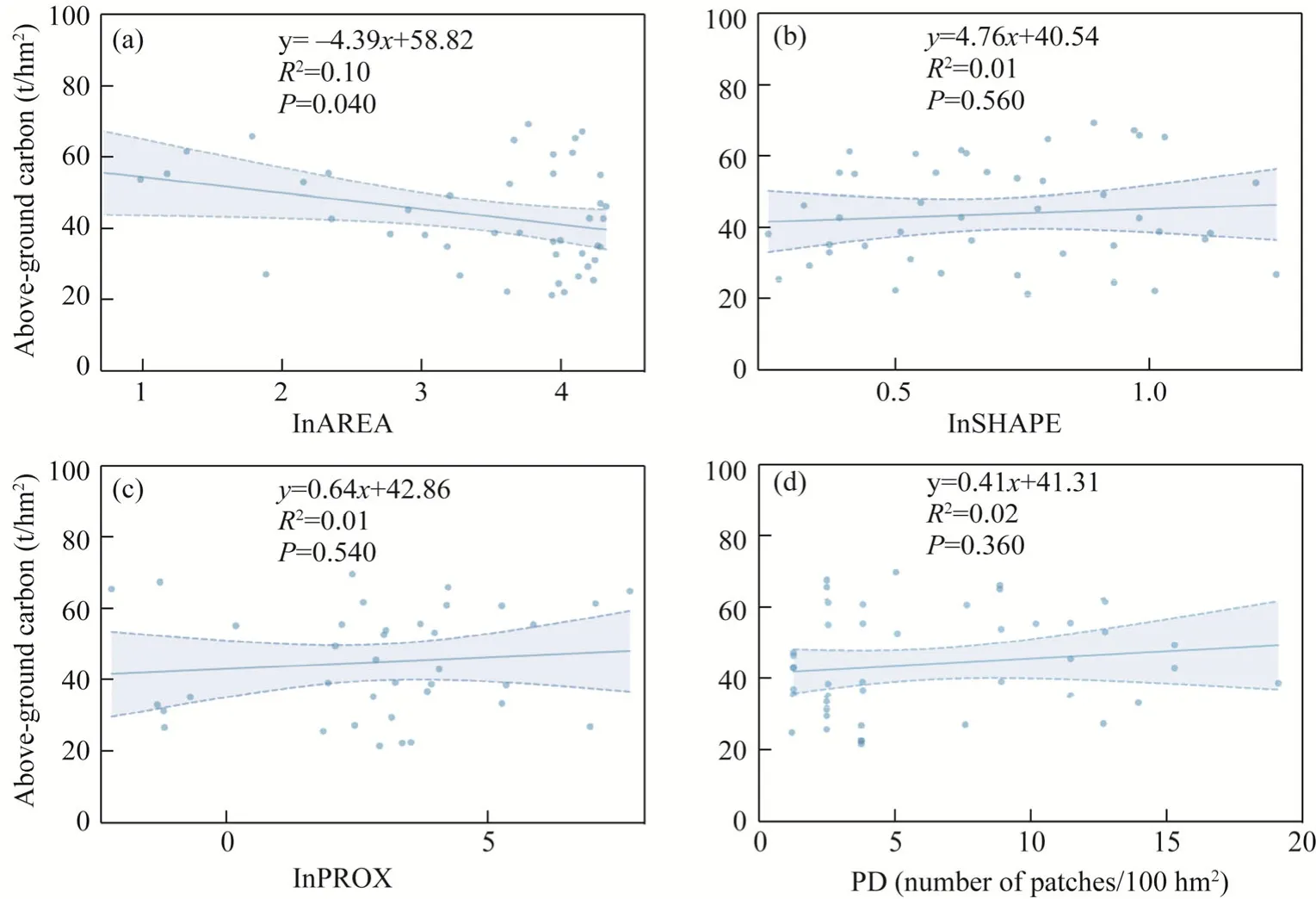
Fig.6 Relationships between four types of fragmentation indices and carbon storage.(a), AREA (patch area);(b), SHAPE (edge); (c), PROX (isolation); (d), PD (patch density).Shaded area represents the 95% confidence interval of the regression.
3.4 Relationships between four types of fragmentation indices and tree density
There was a significant positive correlation between AREA and tree density (Fig.7a), and SHAPE and PD had significant negative effects on tree density, indicating that tree density decreased significantly with increasing fragmentation levels (Fig.7b-d).
3.5 Determinant of forest carbon storage
SEM analysis explained 36% of the variation in AGC (Fig.8).Pvalue of the chi-square test was less than 0.05, meaning a good model fit.In stand structure, tree density had no significant effect on carbon storage, but canopy coverage had a significant negative effect on carbon storage(P<0.010).TWI had no significant effects on carbon storage, canopy coverage, or tree density.DTE had no direct effect on carbon storage, but indirectly negatively affected by canopy cover(P=0.040).Compressed fragmentation could not only directly and positively affect carbon storage(P<0.010), but also indirectly and negatively affect carbon storage by canopy coverage (P<0.010),suggesting that fragmentation is an important influencing factor on forest carbon storage.
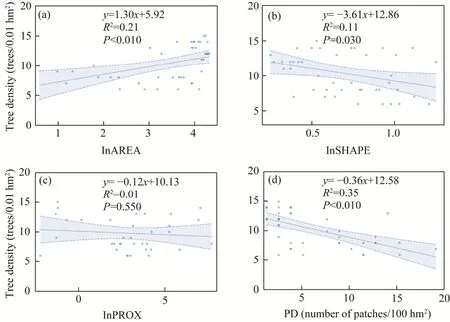
Fig.7 Relationships between four types of fragmentation indices and tree density.(a), AREA (patch area); (b),SHAPE (edge); (c), PROX (isolation); (d), PD (patch density).Shaded area represents the 95% confidence interval of the regression.
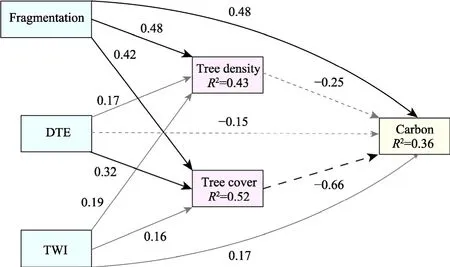
Fig.8 Structural equation model for combined effects of fragmentation, edge effect, abiotic factor, and stand structure on above-ground carbon in plantation forests on the Loess Plateau.Solid lines indicate positive impacts,dashed lines indicate negative impacts, and grey lines indicate coefficients that were not significant at P>0.050 level.DTE, distance to edge; TWI, topographic wetness index.
4 Discussion
4.1 Combined factors affecting above-ground carbon storage
The results from this study suggested that fragmentation can either affect carbon storage through direct effects or, together with DTE, indirectly and negatively affect carbon storage by influencing canopy coverage (Fig.8).The absence of a significant direct effect of tree density on carbon storage challenges has established a consensus (Sanchez and Lindsell, 2017; Khan et al., 2020).Our findings corroborated the significant and negative impact of canopy coverage on carbon storage.Dense canopy cover reduces light, which affects the understory canopy and subsequent carbon sequestration (Ohara and Ushimaru, 2015).Trees at lower ecological niches in the forest floor receive fewer light resources (Fletcher, 2005), and the intensity of light received through the blockage of the upper canopy diminishes, resulting in less intensive photosynthesis and accumulation of less biomass (Rambo and North, 2009).As with the height and DBH data collected in continuous and fragmented forests (Fig.5), the presence of higher ecological niche of black locust in continuous forests affects the overall carbon storage (Fig.4).In previous studies,topography has been recognised as an important factor because it can affect water storage and soil moisture (Sfair et al., 2016), but our investigation challenges this perspective, due to the scarcity of rainfall in the Ansai District, and the limited amount of water that can be stored in the terrain leads to a lack of clear distinction between the lowlands and the highlands, thus the role of the terrain humidity factor is not significant (Li and Liu, 2014).Although DTE exhibited no direct influence on carbon storage, our findings align with the principles of edge effects.The indirect negative effect indicates that proximity to forest edges can induce alterations in microclimates and light regimes, subsequently shaping canopy structure and affecting carbon storage (Sfair et al.,2016).The direct positive effect of the compressed fragmentation factor on carbon storage is consistent with the other studies (Socolar et al., 2016; Jin et al., 2021), highlighting the positive influence of landscape heterogeneity on carbon sequestration.Diverse types of forests accommodate various vegetation types, thereby contributing to elevated carbon storage level(Krishnadas et al., 2018).Additionally, the indirect negative effect is due to its positive impact on the canopy coverage (Ries et al., 2017).This phenomenon could be attributed to the constrained availability of localized resources within fragmented forests, posing challenges in developing a dense canopy due to the dispersed distribution of trees engaged in competitive interactions(Nadkarni, 1994; Hirose, 2005).Conversely, in continuous forests, the heightened competitiveness among trees is more probable, contributing to the establishment of a more compact canopy structure (Hirtreiter and Potts, 2012; Tarvainen et al., 2015).
4.2 Fragmentation affecting above-ground carbon storage
Contrary to our previous hypothesis, AGC storage in fragmented forests was nearly 50.0% higher than that in continuous forests (Fig.4).This result attributed to differences in stand structure characteristics and the proportion of large trees between continuous and fragmented forests.In the present study, the average DBH of black locusts in fragmented forests was significantly higher than that in continuous forests; only 4.2% of trees in continuous forests had a DBH >20 cm,whereas nearly 50.0% of trees were in fragmented forests.Large trees play an important role in carbon storage in fragmented forests.In contrast to natural forests, carbon storage in fragmented forests is lower than that in the continuous forests (Osuri et al., 2014; Benítez-Malvido et al.,2017; Islam et al., 2017; Rolo et al., 2018).The result can be attribute to the differences in species composition and biodiversity between natural and plantation forests.Natural forests can accommodate a high number of species, and this diversity promotes the effective utilization of resources and increases the possibility of species with high functional properties in the community (Cardinale et al., 2012; Rolo et al., 2018).In contrast, plantations rely on a single species to fix carbon, highly similar species composition and high tree density of planted forests,which can lead to intense competition for specific soil nutrients (Gao and Wang, 2018; Sandoval López et al., 2018).According to Liebigs' Law of the Minimum, the scarcity of certain soil nutrients may limit tree growth in continuous forests (Stine and Huybers, 2017).
Among the four landscape indices, AREA had a significant negative effect on carbon storage in black locust forests (Fig.6), which may have been caused by insufficient growth resources and high-intensity growth competition (Cardinale et al., 2012).The larger the patch, the higher the survival rate of black locusts (Fig.6), and thus, the greater the competition for resources.Therefore, light and other resources limit the growth and development of black locusts in large patches.The positive correlation between larger patch size and increased tree survival rate is in agreement with fundamental ecological principles (Kohyama, 2006; del-Val et al., 2007).The heightened biodiversity and establishment of symbiotic relationships within larger patches significantly contribute to the overall well-being of trees (Haddad et al., 2015).Favourable conditions that including soil nutrient and exposure to sunlight, are prevalent in larger patches,creating an optimal environment for tree growth (Jacodziński and Oleksyn, 2009).The optimal environment, in turn, protects trees against environmental stress (Bianchi and Kersten, 2014).Additionally, the regulated microclimates within larger patches, facilitated by augmented vegetation coverage, act as a protective shield for trees, shielding them from extreme fluctuation and providing a conducive setting for tree establishment and long-term survival (Harper et al.,2015).
4.3 Effect of fragmentation on tree density
A regression plot of the four indices characterising fragmentation (AREA, SHAPE, PROX, and PD) versus tree density indicated that tree density decreased as fragmentation increased,indicating lower tree survival (Fig.7).Black locust forests were planted in 1999, and the initial planting density was consistent (Lü et al., 2017).However, this study showed that tree density in fragmented forests was lower than that in continuous forests, which is consistent with our previous hypothesis.This result may be caused by drought in Yan'an City, where the main meteorological disasters are drought, low-temperature frost, and dry and hot wind, with an annual average wind speed of 1.7 m/s.With the dry, high-temperature, and arid climatic conditions in Yan'an City, slow winds can remove water from the trees (Laurance et al., 2002).In black locusts,drought is likely to cause death in fragmented forests.However, in continuous forests,microclimate resources such as water, temperature, and humidity are more readily available(Camille et al., 2020), and the survival rate of trees is higher (Fig.6).Our study aligns with existing research (Püttker et al., 2020; Pöpperl and Seidl, 2021), highlighting the significance of canopy structural diversity and resource availability in shaping the observed robust survival patterns in continuous forests.The enhanced landscape connectivity within these ecosystems not only fosters symbiotic relationships but also establishes a web of ecological interaction that fortifies the overall health and vitality of trees, creating conditions conducive to sustained survival (Hedwall et al., 2019).Additionally, the resource-rich environment, characterised by abundant soil nutrients and optimal sunlight exposure, plays a pivotal role in creating an optimal setting for tree growth, thereby enhancing their capacity to withstand environmental stress (Erdős et al., 2018; Krishnadas et al., 2018).
4.4 Relationship between tree height and diameter
When DBH of black locust was >10 cm, given the same DBH, the number of trees in continuous forests was higher than that in fragmented forests.Because of the higher tree density, the intensity of competition for light, water, and soil nutrients in continuous forests was higher than that in fragmented forests.Competition results in the height of black locusts in the continuous forests forming a gradient; only a few individuals have growth advantages, and the growth of black locusts in forests with a lower tree density is faster (Li et al., 2008).When DBH of black locusts in the two types of forests reached approximately 10 cm, to compete for light resources, black locusts in continuous forests adopted the growth strategy of distributing nutrients for vertical growth (Holbrook and Putz, 1989).However, it was more inclined to distribute nutrients for horizontal growth to enhance resistance to environmental stress in fragmented forests (Eldpausch et al., 2011).The wind stress on trees in fragmented forests increases the probability of tree destruction (Laurance et al., 1997).
4.5 Future carbon storage trends of continuous forests
Plantations of tree species with high wood density can exceed the carbon storage capacity of natural forests at harvest age (Bonner et al., 2013; Yang et al., 2019a), but due to the regular harvesting of commercial forests, plantation forests are considered ineffective for long-term carbon storage (Zaninovich and Gatti, 2020).However, the non-commercial nature of plantations on the Loess Plateau is of great significance for long-term carbon storage.A previous biomass simulation study of black locust forests with different densities on the Loess Plateau showed that the most suitable planting density of black locusts in Yan'an City was 15 trees/0.01 hm2, which was equivalent to continuous forests in this study, and the highest biomass was obtained during the growth of black locust (Li et al., 2008).The black locustforests measured are 20 a, which is not yet a period of carbon sequestration dominance in continuous forests.Some studies have shown that black locust forests with a density of 15 trees/0.01 hm2have the highest carbon sequestration compared with the other density of black locustforests around 30 a (Shahid and Joshi, 2018; Cabrera-Ariza et al., 2021).
Our results demonstrated a phenomenon consistent with similar studies in temperate forests(Stine and Huybers, 2017; Sandoval López et al., 2018), where carbon storage was higher in the pre-fragmented forests than in continuous forests.This observation is consistent with previous studies showing that in the early stage (Laurance et al., 2002), fragmented forests may exhibit higher levels of carbon storage due to lower tree densities, more diverse spatial distributions, and more abundant growth resources (Capellesso et al., 2020; Li et al., 2020).However, some results also highlight the interesting phenomenon that tree growth rates stabilise over time, whereas continuous forests gradually accumulate higher levels of carbon storage (Deng et al., 2014).Research showed that with the trees mature in continuous forests as stand ages, their competition for resources tends to decrease, resulting in increased individual tree growth and carbon accumulation (Li et al., 2014; Jagodzinski et al., 2019).Continuous forests have greater tree densities that give them an advantage in carbon fixation (Yang et al., 2019b).Additionally, the development of more complex structural features in older continuous forests (Rolo et al., 2018),such as larger trees and diverse canopy structural diversity, may contribute to higher levels of carbon storage (Rambo and North, 2009).These structural attributes are known to positively influence carbon sequestration, which is consistent with previous findings (Rolo et al., 2018;Sandoval López et al., 2018).
According to the relationship between the growth law and current allometric growth of black locusts (Fig.5), we predicted that the carbon storage of continuous forests will exceed that of fragmented forests in the later growth stages.This prediction depends on whether the subsequent growth of black locust forest maintains this height-diameter growth relationship, which requires a long-term observation.
5 Conclusions
Fragmentation had several effects on the tree density, plantation structure, and carbon storage.Tree competition for resources not only changed the choice of survival strategies in the two types of forests but also affected the growth and development of trees, thus impacting carbon storage.Although evidence from our study suggested that carbon stocks were currently higher in fragmented forests than in continuous forests and that AREA had a significant negative effect on carbon stocks at current stage, however carbon storage and ecological benefits of continuous forests in the later period exceeded those of fragmented forests.Therefore, in the follow-up tree planting, the influence of patch area, edge, and patchy connection on plantation should be investigated, and policymakers can incorporate the following measures into plantation to compensate for the loss of carbon storage potential due to fragmentation: (1) expanding the area of plantation patches; (2) repairing the edges of complex-shaped patches; (3) enhancing the connectivity of similar patches; and (4) applying specific nutrients to different plantations at regular intervals.
Conflicts of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (32201429) and the College of Landscape Architecture and Art, Northwest A&F University, China.
Author contributions
Conceptualisation: LI Xian, LEI Hangyu; Methodology: LI Xian, LEI Hangyu; Formal analysis: LEI Hangyu;Writing - original draft preparation: LEI Hangyu; Writing - review and editing: LI Xian; Funding acquisition: LI Xian; Data curation: DUAN Dantong; Investigation: DUAN Dantong, CHEN Yi, GUO Huifeng, LI Jiangtao, LEI Hangyu; Project administration: LI Xian; Resources: LI Xian; Software: LEI Hangyu; Supervision: LI Xian;Validation: LEI Hangyu; Visualisation: LEI Hangyu.All authors approved the manuscript.
Appendix

Table S1 Parameter statistics for structural equation modelling

- Journal of Arid Land的其它文章
- Exploring groundwater quality in semi-arid areas of Algeria: Impacts on potable water supply and agricultural sustainability
- Runoff change in the Yellow River Basin of China from 1960 to 2020 and its driving factors
- A CMIP6-based assessment of regional climate change in the Chinese Tianshan Mountains
- Influence of varied drought types on soil conservation service within the framework of climate change:insights from the Jinghe River Basin, China
- Land use and cover change and influencing factor analysis in the Shiyang River Basin, China
- Effects of drip and flood irrigation on carbon dioxide exchange and crop growth in the maize ecosystem in the Hetao Irrigation District, China