
光照强度对大口黑鲈游泳协作能力的影响
2024-02-07 07:29吕义淞文彦慈杭晟煜倪伟强蔡海莺叶章颖
水生生物学报 2024年2期
吕义淞 赵 建 文彦慈 杭晟煜 倪伟强 蔡海莺 叶章颖,
(1.浙江科技学院生物与化学工程学院, 杭州 310023; 2.浙江大学生物系统工程与食品科学学院, 杭州 310058;3.浙江大学海洋研究院, 舟山 316021)
我国是水产养殖大国, 养殖产量占全球总量60%以上, 且已远超捕捞渔业所提供的水产品产量[1]。就水产养殖而言, 环境调控至关重要, 直接影响水产品的质量和产量, 尤其是在工厂化循环水养殖(Recirculating aquaculture system, RAS)模式下[2—5]。光照是影响鱼类生长的重要环境因子, 是RAS中调控的重点, 主要通过光色、光周期和光强作用于养殖对象[6—9]。不同光色对鱼类应激水平和生长率等生理指标产生不同的影响, 如: 处于黄光和蓝光下短须裂腹鱼(Schizthrax wangchiachii)的谷丙转氨酶含量相对自然光照下较高, 而皮质醇的含量在蓝光条件下显著高于自然光和黄光[10]; 红光使克氏海葵鱼(Amphiprion clarkii)体内的褪黑激素合成酶活力高于其他光色环境, 从而增强了褪黑激素的分泌以及氧化应激反应, 最终起到保护鱼体的作用[11]。光周期通过调节鱼类的内源性节律, 对养殖鱼类的生长、发育和繁殖具有一定影响, 如: 长光周期加快雌性光唇鱼(Acrossoeheilus fasciatus)的卵巢发育且提高其血浆褪黑素含量[12]; 在较长光周期(16L∶8D)养殖的黄金鲈(Perca flavescens)总体重净增量是自然光照下的2.90倍, 且饵料系数相对于自然光照条件下降低54.8%[13]。光照强度的改变则会影响幼鱼的成活率以及发育速率。当光照强度较弱时(<500 lx), 克氏双锯鱼(Amphiprion clarkii)幼鱼的发育速率、成活率与光照强度呈正相关, 当光照强度达到2000 lx时其发育速率显著下降[14]。类似的现象在褐石斑鱼(Epinephelus bruneus)和豹纹鳃棘鲈(Plectropomus leopardus)中也被发现, 当光照强度较弱时(1—500 lx), 随着光照强度的增强, 其幼鱼成活率逐渐增加[15,16]。
光照不仅影响着鱼类的生理状态, 还作用在其行为上。如短须裂腹鱼(Schizothorax wangchiachi)容易被黄光、红光和绿光吸引, 而对蓝光选择逃避,且当其处在22.6—39.5 lx光强范围内时, 被光照吸引的总尾数与光照强度呈正相关; 当处在50.1—64.7 lx光强范围内时则为负相关[10]。瓦氏黄颡鱼(Pelteobagrus vachelli)对光色不敏感, 但更容易被0—10 lx的光照吸引(该光色区域内的出现次数百分比: 78.33%, 其余光照下分别为10.00%、7.33%和4.33%); 不同光照强度对鲢(Hypophthalmichthysmolitrix)的吸引没有显著性差异, 但蓝光更能吸引鲢(该光色区域内的出现次数百分比35.67%, 其余分别为12.67%、30.33%和21.33%)[17]。红鲫(Carassius auratus)对灯光颜色的喜好顺序为黑色>蓝色>红色>绿色>橙色>黄色, 且该品种对空间位置优先择取鱼缸的左右两侧[18]。当前光照对鱼类行为影响的研究主要集中在对养殖品种个体趋光性(光强和光色)的探究上, 少有关注鱼类群体行为, 尤其是其游泳协作能力。
游泳能耗是鱼类日常能量支出中的主要部分,最高可达40%[19,20], 在很大程度上影响着养殖效益。鱼群内部的游泳协作可使群内个体相对于单独游泳时的游泳能耗降低(理论上或可高达70%[21]),避免不必要的能量损耗[22,23]。研究表明, 环境因素直接影响着鱼类游泳协作能力。Hang等[24]发现在工业化水下噪声的影响下, 大口黑鲈的群体结构较为松散, 具体表现为相邻鱼之间的角度、距离增大;Paolo等[25]发现低氧状态时, 为保证个体鱼能够尽可能多地从低氧水体中获取氧气, 鱼群中个体之间的相对距离增大; Cooper等[26]发现当水温分布不均匀时, 具有“偏好温度(Preferred temperatures)”的个体更倾向于停留在其偏爱的温度区域, 而选择不跟随鱼群; 流速则会导致鱼群的阵型配置发生改变,高流速下的鱼群更倾向于采取并排排列的阵型, 而非低流速下的“钻石(Diamond-shaped)”阵型[23]; 此外, Armstrong等[27]发现暴露在油污中会使鱼群中的个体平均移动速度降低及相邻个体之间的距离增大, 从而削弱了鱼群的凝聚力(Cohesion)。然而,光照强度对鱼群游泳协作能力的影响还鲜有报道。
大口黑鲈由于经济价值高, 国内淡水养殖热门品种, 被誉为继“四大家鱼”后的第五大家鱼。本研究以大口黑鲈为研究对象, 探究不同光照强度对循环水养殖下鱼类游泳协作能力的影响, 旨在为循环水养殖大口黑鲈光环境调控提供理论依据。
1 材料与方法
1.1 实验材料
大口黑鲈于2022年11月购自杭州唯康农业开发有限公司, 总数为150尾, 购买后置于本实验室规格为高120 cm×直径100 cm的循环水圆形池内驯养15d。驯化期间每天上午10: 00以通威商业颗粒饲料饱足投喂1次, 投喂1h后立即清除残饵和粪便, 日换水量约为驯化水体的25%。水温控制在(23±1)℃并保持24h不断曝气, 水体溶氧水平为(6.2±0.5) mg/L, 光周期为自然光照周期(12L∶12D)。在驯化完毕后选择个体健康且大小相近大口黑鲈[体重(25±5) g,体长(15±2 cm)]共66条作为实验对象。
1.2 实验系统
本实验系统主要由3部分组成: Brett-type游泳隧道[28]、摄像录制系统及照明系统。游泳隧道的主要作用是提供一个流速稳定可控的实验环境, 摄像录制系统用于视频数据的录制、存储与分析, 照明系统用于提供实验光源。
游泳隧道(图1)主要由转速可控的电机和内、外水槽(材料: 透明亚克力)构成。电机通过频率控制器改变频率从而达到对其转速的精准控制, 电机输出端为直径30 cm的螺旋桨, 通过螺旋桨的转动在内水槽中产生稳定流速的非湍流水体[29], 水体温度保持在22—24℃, 流速为0.7 BL/s (体长/s)。内水槽(长180 cm×宽80 cm×高40 cm)开设长100 cm×宽30 cm×高30 cm的游泳槽, 游泳槽内水体与实验对象直接接触。外水槽(长220 cm×宽110 cm×高40 cm)的主要作用保证内外水槽液面高度一致(不存在压强差), 从而确保游泳槽内的流速恒定及整个Bretttype游泳隧道的稳定运转。

图1 Brett-type游泳隧道结构示意图Fig.1 Schematic diagram of the structure of the Brett-type swimming tunnel
摄像录制系统主要由相机(BASLER acA2040-90 μm, 分辨率: 2448×680, 帧率: 60 fps)、刻录系统构成, 相机架设于实验对象所在游泳槽的中央正上方, 并通过Basler程序进行对焦和录制画面的参数调试。
照明系统由红外补光灯和LED灯构成。红外补光灯(功率: 90 W, 长22.5 cm×宽18.5 cm×高15 cm。)的作用是给黑暗条件下的实验环境提供摄像机可见光源, 红外补光灯架设在高度、角度可灵活调节并锁止的相机支架上。LED灯管(功率: 0—35W可调, 长118.5 cm×宽2.1 cm×高3.2 cm)通过底部固定、高度相同且角度可调节并锁止的灯管夹平行架设在实验对象所处游泳槽正上方, 以保证游泳区域内光照相对匀称。光照强度检测通过光照强度测试仪(希玛AS823)在游泳槽注满水后由浅至深测量。
1.3 实验方法
实验开始前, 先挑选一对体型大小接近的实验对象放入实验室内规格为长30 cm×宽35 cm×高45 cm的暂养水箱[水温(23±1)℃]中饥饿处理48h, 以排空肠内残余消化物, 避免特殊动力作用对实验对象耗氧率或游泳能力的影响[30]。
具体操作步骤: (1)设置好实验灯光(每组鱼依次经历0、300—500、2200—2500和3300—3900 lx的光强环境), 并在禁食处理结束后将两条鱼用抄网轻轻放入游泳槽[每组实验前将曝气24h、温度(23±1)℃溶氧水注入该游泳隧道]。(2)给予实验对象30min休息时间, 之后通过频率控制器调整输出频率, 控制电机在2min内缓慢提升隧道内流速至0.7 BL/s, 并在该流速下适应5min后通过置于游泳隧道正上方的摄像机拍摄记录(每个记录周期: 记录3min+停止记录2min, 每组实验对象共记录3个周期)。(3)在每种灯光实验结束后, 将流速在2min内缓缓调节至0, 并关闭灯光以供实验对象在黑暗中休息30min。(4)在休息结束后, 改变灯光条件重复实验步骤(1)、(2)、(3)、(4)直至每组大口黑鲈经历完所有实验灯光。在本实验中, 每组实验结束后重新更换游泳隧道中的水体, 共计有33组实验对象完成本次实验。
1.4 数据分析与处理
游泳能耗量化指标鱼类在一般流速(< 3 BL/s)下的游泳能耗来自有氧代谢产生的能量, 因此耗氧量反映了其游泳时的能量支出[31]。已有研究表明鱼类的摆尾频率与其耗氧量的变化呈正相关[32—34],故本实验采用matlab计算不同光照强度下大口黑鲈的平均摆尾频率, 以代表其游泳能耗(图2)。平均摆尾频率计算公式[32]如下:

图2 摆尾频率量化方法示意图Fig.2 Schematic diagram of the tail beat frequency quantization method
式中,NT为个体鱼在稳定游泳时的摆尾周期数,t为稳定摆尾总时间。
游泳协作能力量化在研究鱼类行为的相关报道中, 常常采用量化个体鱼之间的相对距离和位置情况来评价鱼群的整体游泳协作能力[24,29,35]。本次实验通过matlab计算不同光照强度下两条鱼之间的相对距离与相对位置分布频率, 并绘制热度图[36]。相关计算公式如下:
A.相对距离(欧氏距离)
式中,x1与y1为焦点鱼在摄像机俯视镜头拍摄得到的二维平面下的横坐标与纵坐标, 同理x2与y2为邻近鱼的横坐标与纵坐标。
B.相对位置分布频率
式中,F1,2,3,4为各个位置下邻近鱼相对焦点鱼所处位置的帧数总和(1、2、3和4分别代表前、后、左和右),Ft为总帧数。焦点鱼的前、后、左和右的方位划分方法: 以其头部为原点建立二维坐标系后每45°、135°、225°和315°为分界线, 每90°作为一块方位所属区域。具体划分示意图3。
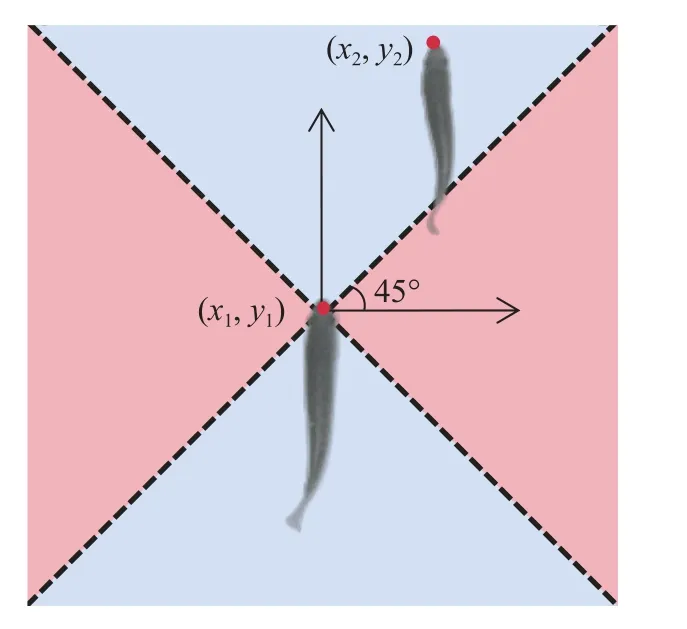
图3 焦点鱼周边方位示意图Fig.3 Schematic diagram of the perimeter of the focal fish
C.攻击次数
由于大口黑鲈具有一定的攻击性, 且在本次实验中攻击行为的出现较为频繁, 在一定程度上影响了个体间的游泳协作, 因此本研究统计了不同光照强度下鱼群攻击行为(包含追逐、撕咬和威胁)的发生次数, 以促进鱼群游泳协作能力的深入分析[37]。
数据统计和分析本研究利用SPSS单因素重复测量方差分析(ANOVA, Repeated measures analysis)对数据进行统计和分析, 并通过最小显著性差异法(LSD)进行事后检验。统计值使用平均值±标准差(mean±SD)表示,P<0.05表示差异显著。
2 结果
2.1 不同光照强度下大口黑鲈鱼群的游泳能耗
大口黑鲈在光照强度分别为0、300—500、2200—2500和3300—3900 lx下的摆尾频率分别为(1.73±0.28)、(1.84±0.37)、(2.03±0.38)和(1.98±0.30) Hz (图4)。结果显示: 在光强为0时, 鱼群的游泳能耗显著低于其他实验组(P<0.05); 而随着光照强度的增加, 鱼群游泳能耗呈现出上升趋势且各组间差异显著(P<0.05), 直到光强上升至3300—3900 lx时鱼的游泳能耗相对2200—2500 lx组出现了下降,但差异并不显著(P>0.05)。
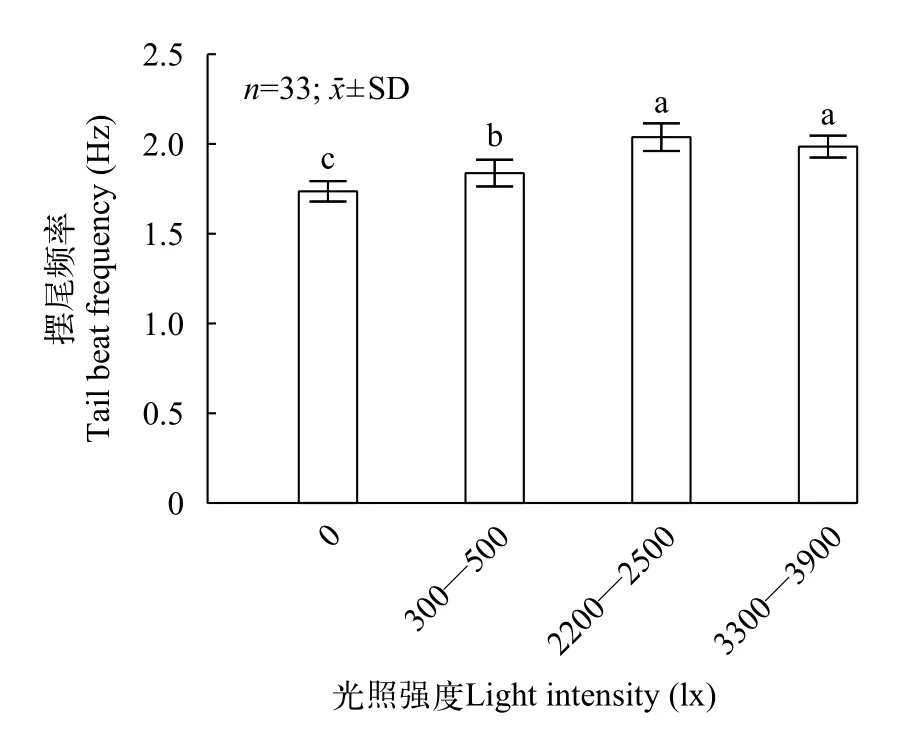
图4 不同光照强度下大口黑鲈的摆尾频率Fig.4 The tail beat frequency of largemouth bass under different light intensity
2.2 不同光照强度下大口黑鲈鱼群的游泳协作能力
光照强度对大口黑鲈的相对距离有显著影响(P<0.05), 虽然2200—2500 lx组的相对距离与300—500 lx组和3300—3900 lx组无显著差异(P>0.05), 但是总体上两组间的相对距离呈现出随着光强的增强而增大的趋势(图5), 同时在热度图中也可观察到当光强逐步增大时两鱼间距离的扩散变化过程(图6)。大口黑鲈在光照强度不断增强的光环境下更倾向于采取前后排列的位置分布(图5),且该位置分布的出现频率在各组间存在显著差异(P<0.05)。热度图(图6)反映了邻近鱼相对于焦点鱼的具体空间位置分布情况。
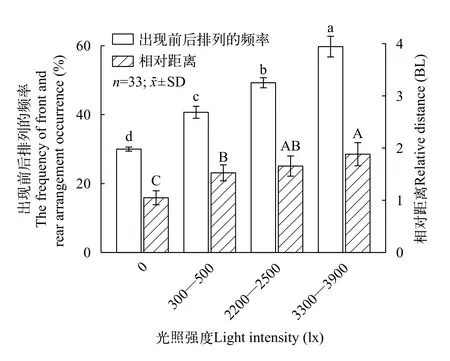
图5 大口黑鲈在不同光照强度下的相对距离及出现前后排列的频率Fig.5 Relative distance and the frequency of front and rear arrangement occurrence of largemouth bass under different light intensity

图6 相对位置分布热度图Fig.6 The heatmap of relative position distribution
2.3 不同光照强度下大口黑鲈鱼群攻击次数
不同光照强度(分别为0、300—500、2200—2500和3300—3900 lx)下大口黑鲈鱼群攻击次数分别为(2.39±0.629)、(3.79±0.76)、(6.73±1.36)和(7.06±1.62)次/180s, 其中, 3300—3900和2200—2500 lx下发生的攻击次数最高且与0组差异显著(P<0.05), 略高于300—500 lx组, 但差异不显著(P>0.05)。总体呈现出攻击次数随光照的增强而逐步增大。
3 讨论
鱼群个体间相对距离和空间位置选择影响着鱼群游泳协作能力的表达, 并继而决定了鱼群的游泳能耗[32,38]。鱼群个体间的距离和空间位置的选择由许多因素决定, 如领导力、运动能力及饥饿程度[35,39,40]。本实验发现在黑暗条件下两鱼相对距离最短且很少采用前后排列(图5), 这可能是因为在短距离、左右排列的条件下两鱼可更好地利用彼此摆尾时产生的水动力学收益(Hydrodynamic benefits), 从而节省游泳能耗[22,41](图4)。此外在黑暗条件下, 两鱼间出现的攻击次数较少(图7), 在多数情况下采用安静、平稳的协作式游泳, 这可能是为了减少自身受到捕食的风险并增加生存的机会[42]。
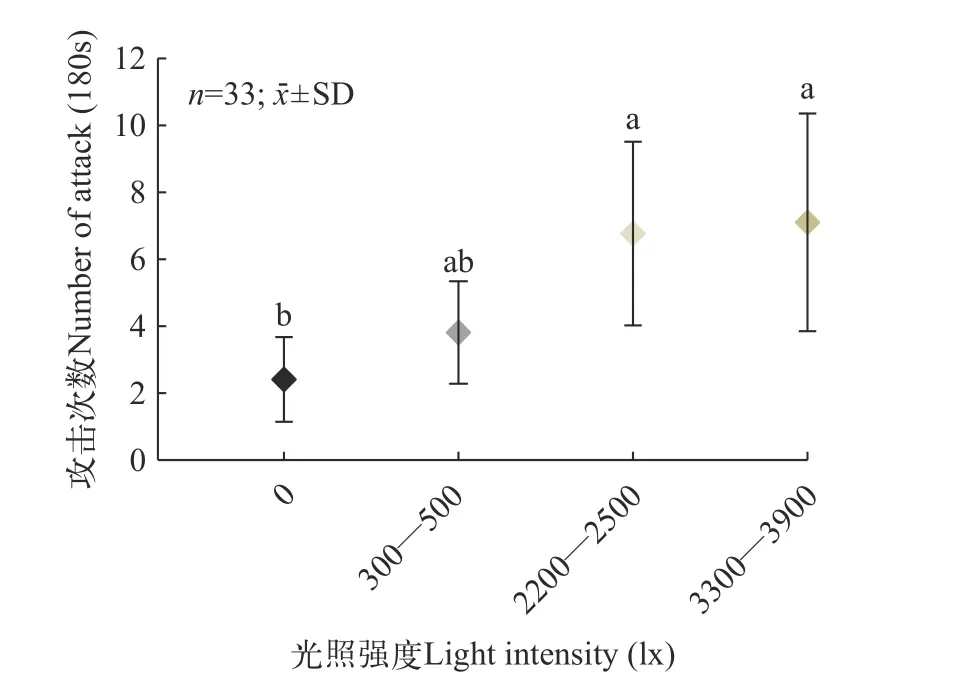
图7 不同光照强度下的攻击次数Fig.7 The number of attack under different light intensity
研究发现, 当光照增强时, 两鱼表现为独立趋光探索、相互追逐的活动增强, 协作游泳较少, 具体体现在个体间的相对距离增大、更多地采取前后分布排列(图5)及攻击现象加剧(图7), 同时伴随着高游泳能耗(图4)。这一现象的出现可能是由于光强的增大导致了鱼群个体生理应激反应的增强,从而使得个体游泳时的活跃度及耗氧量增强[43]。类似现象也在其他研究中被报道, 如: Schleuter等[44]发现欧亚鲈(Perca fluviatilis)在白天正午时的活跃度与傍晚相比提高了69.3%, 正午时[489 mg O2/(kg·h)]常规代谢率(Routine metabolic rate)相对于傍晚时[220 mg O2/(kg·h)]提升了的1.22倍; 而梅花鲈(Gymnocephalus cernuus)正午时的活跃度相对傍晚时提高了61%, RMR则从176提升到289 mg O2/(kg·h), 约为64%; 正午时大菱鲆(Scophthalmus maximus)的耗氧量[248 mg O2/(kg·h)]显著高于早晨[162 mg O2/(kg·h)]和傍晚[171 mg O2/(kg·h)][45]。值得一提的是, 本研究中的最大光照强度3300—3900 lx对应的摆尾频率相较于2200—2500 lx有轻微下降但不明显(P>0.05)。一般来说, 对鱼类生长无明显影响的光照强度阈值为600—2000 lx, 当光照过强时, 鱼类会展现出游泳疲劳和死亡率上升的现象[46,47], 比如海鲈(Dicentrarchus labrax)、露斯塔野鲮(Labeo rohita)和团头鲂(Megalobrama amblycephala)[47—49], 这可能是本次实验中最高光强3300—3900 lx下摆尾频率相对2200—2500 lx组出现不显著下降的原因。
事实上, 光照的增强还会提高鱼群好斗性[37,50],这可能也是制约鱼群游泳协作能力表达的重要因素。实验中观察到攻击现象的发生会导致两鱼间相对安静的平稳游泳状态被破坏, 主要表现为个体对另一个体的撕咬、追逐及威胁(用嘴轻轻触碰另一个体的身体), 进而导致两鱼长时间保持较远的相对距离, 展现出彼此提防、警戒的现象, 同时这也有可能是由于过强的光照会对群体内个体的视觉感知产生干扰, 导致个体难以识别周围的伙伴或敌人而保持较远的相对距离[51,52]。此外, 主动发起攻击的个体更经常地出现在群体前端, 考虑到每条实验对象都经历了2d的禁食, 因此主动发起攻击的个体可能更想通过占据前端获取优先进食的机会[53,54]。而被攻击的个体由于受到了频繁的攻击,更倾向于躲藏和减少活动, 在本实验中观察到被攻击的个体一般会躲藏在攻击者的后端, 且试图依赖水流方向拉开与攻击者间的距离, 原因可能是为了减少暴露在攻击者的视野范围内, 以降低再次被攻击的可能性。
本研究表明, 当光照强度较低时鱼群游泳协作能力较好, 游泳能耗较低。而当光照增强时, 鱼群游泳协作能力由于光强刺激、同类间攻击现象加剧而受到制约, 因此, 在实际生产中大口黑鲈养殖可更多地选择光强较弱(0—500 lx)的养殖环境, 以避免由于光照过强导致的鱼群游泳协作能力下降,降低鱼群游泳能耗及攻击现象的发生频次, 减少生产养殖成本。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
基层中医药(2021年6期)2021-11-02
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
创新作文(小学版)(2018年10期)2018-08-20
中外文摘(2017年19期)2017-10-10
广东饲料(2016年7期)2016-12-01
电测与仪表(2016年3期)2016-04-12
电测与仪表(2016年20期)2016-04-11
爆笑show(2015年8期)2015-09-22
