
饲料中小麦胚芽对黄颡鱼雌性亲鱼繁殖性能的影响
2024-02-07 07:29李亚宁樊启学沈志刚
水生生物学报 2024年2期
李亚宁 陈 敏 刘 洋 韩 磊, 何 焱 樊启学, 沈志刚
(1.华中农业大学水产学院, 农业农村部淡水生物繁育重点实验室, 教育部长江经济带大宗水生生物产业绿色发展工程研究中心, 池塘健康养殖湖北省工程实验室, 武汉 430070; 2.湖北黄优源渔业发展有限公司, 武汉 430299)
水产养殖业健康可持续发展最关键的前提是充足稳定的苗种供应, 而苗种的数量和质量首先取决于具有优良品质、性腺发育良好的亲鱼。黄颡鱼(Tachysurus fulvidraco)是一种中小型、杂食性的经济鱼类, 主要在我国内陆水体中的底层生活。因其肉质细腻爽口、无肌间刺和营养丰富等优点, 已成为我国最重要的特色淡水鱼类之一[1,2]。养殖覆盖全国28个省份, 有全民消费趋势。中国渔业统计年鉴显示, 黄颡鱼年产量从2011的21.7万吨增长到2021年58.8万吨[3], 产业的高速发展得益于人工繁殖技术的突破及养殖技术的成熟, 更因为性状优良的新品种“全雄1号”和杂交黄颡鱼“黄优1号”(黄颡鱼T.fulvidraco♀×瓦氏黄颡鱼T.vachelli♂)的成功推广。由于雌性黄颡鱼生长速度显著小于雄性或杂交种[4], 经济效益低, 养殖户更倾向于选择饲养全雄或者杂交黄颡鱼。当前杂交黄颡鱼“黄优1号”的成功推广, 推动了全国杂交黄颡鱼养殖的全面普及, 但同时导致雌性亲鱼养殖量的大幅度缩减。此外, 黄颡鱼人工繁殖过程中密集的人工操作不可避免地给亲鱼带来损伤, 引起应激及细菌病毒感染[5],导致雌性亲鱼产后死亡率较高, 一般达30%以上[6],加剧了雌性亲鱼数量缺乏的现状。更为重要的是,雌性亲鱼的损失将严重影响黄颡鱼新品种选育进程和效果[6]。因此, 提高亲鱼培育存活率, 提高繁殖效率, 对黄颡鱼产业可持续发展具有重大意义。
小麦胚芽是麦粒最里面的器官, 占据整个麦粒的2.5%左右[7], 通常作为面粉工业中的副产物, 其含有约27%—30%蛋白质, 10%油脂, 47%碳水化合物, 及多种生物活性物质, 如谷胱甘肽、凝集素、黄酮类化合物和植物甾醇等[7,8], 还富含丰富的钙、钾、镁和铁等多种矿物质和微量元素, 具有预防癌症、提高抗氧化及免疫性能、有益生殖系统等功效[8—10]。我国小麦产量居全球第一, 国家统计局数据显示2022年小麦产量1.38亿吨, 较2021年增长0.6%。据相关数据表明, 1000 kg小麦约提取1.1 kg小麦胚芽[11], 其来源稳定、产量充足。近年来, 在雄兔[12]、公猪[13]和南美按实蝇(Wiedemann)[14]中的研究表明, 小麦胚芽可提高动物生殖性能。在水产领域, 朱伟等[15]证明当小麦胚芽替代日本对虾(Penaeus japonicus)饲料中5%—10%鱼粉时能显著提高对虾增重率; 在欧洲鲈(Dicentrarchus labrax)[16]的饲料中添加155 g/kg小麦胚芽可以显著提高饲料利用率, 降低成本。也有研究半滑舌鳎(Cynolossus semilaevis)[17]、杂交石斑鱼(Epinephelus fuscoguttatus♀×Epinephelus lanceolatus♂)[18]等水产动物上探究小麦胚芽作为饲料原料的可行性, 但目前的研究多集中在幼鱼阶段, 在鱼类亲鱼培育及其对繁殖性能影响方面的研究较少, 仅见于几项专利的报道中, 存在巨大空白。
鉴于小麦胚芽营养丰富且具有强大的生理功能, 本研究在黄颡鱼亲鱼饲料中添加适宜含量的小麦胚芽, 分析其对雌性亲鱼肝脏抗氧化性能、血浆生理生化、产前和产后成活率、形体指数及繁殖性能的影响, 探究其作为雌性亲鱼饲料原料的可行性, 为建立科学的雌性亲鱼营养培育模式, 进一步提高全雌黄颡鱼配套系的繁殖性能提供良好的理论基础和科学依据。
1 材料与方法
1.1 实验饲料
实验以添加0、5%、10%、15%和20%(w/w)的小麦胚芽设计5个实验组, 分别设为W0、W5、W10、W15和W20的等氮等脂饲料, W0为对照组,其余为处理组。饲料配方及实测营养组成如表1所示。小麦胚芽购于湖北小笨鱼农业发展有限公司。饲料由湖北小笨鱼农业发展有限公司制作成颗粒料烘干、冷藏于4℃备用。

表1 饲料配方及营养水平 (风干基础)Tab.1 Feed formulation and nutrient levels (air-dried basis)
1.2 实验对象与饲养管理
养殖实验在湖北黄优源渔业发展有限公司室外池塘(面积约2000 m2、水深约1.5—2.0 m) 进行。正式实验前, 将实验鱼在网箱(5 m×5 m×2.8 m)中用W0组饲料暂养1周。
黄颡鱼是分批产卵鱼类, 每年首次产卵后, 经过一段时间培育, 可进行二次甚至多次产卵[19]。华中地区在实际养殖生产过程中, 黄颡鱼雌性亲鱼经过春季营养培育7周左右, 产后再培育4周左右便可达到人工催产的阶段。因此, 投喂实验分两个阶段,即产前培育阶段和产后再培育阶段。产前培育黄颡鱼雌性亲鱼实验从4月17日至6月5日, 养殖培育持续49d。实验鱼来自公司杂交黄颡鱼“黄优1号”雌性亲本的同一亲鱼培育池。实验开始前1天对实验鱼饥饿24h, 挑选体格健壮、规格均匀的个体825尾(79.8±1.4) g, 随机放入5个组15个网箱内(1.7 m×1.7 m×2.8 m), 每组3个平行, 每个平行55尾。每天于17:20—18:40表观饱食投喂1次(体质量的0.2%—1.5%), 并根据实验鱼的摄食情况以及水温等环境因素进行相应调整。实验期间水温为21.0—26.9℃。
产后再培育雌性亲鱼实验从6月27日至7月27日, 养殖培育持续30d。实验鱼是来自同一批次刚完成产卵的雌性亲鱼。正式实验开始前1天对实验鱼饥饿24h, 试挑选体格健壮、规格均匀的825尾(56.7±1.0) g, 这个规格的鱼经历繁殖的人工操作后更易受伤, 存活率较低, 所以选用此规格的雌性亲鱼作为产后再培育的实验对象。随机放入5个组15个网箱内, 每组3个平行, 每个平行55尾鱼。每天(18:10—18:30) 表观饱食投喂1次(体质量的1%—2%), 实验期间水温为28.0—31.6℃。
实验期间溶氧≥5 mg/L, 氨氮<0.2 mg/L, 亚硝酸盐<0.01 mg/L, 光照周期为自然光周期。每天记录水温和死亡数量等。在实验期间的网箱, 用契合形状的钢筋固定以便网箱箱底接触池塘底部, 从而模拟池塘底部环境。
1.3 样品采集
春季产前培育养殖实验结束时, 停食24h, 记录每个网箱实验鱼剩余数量计算成活率(SR)。每个网箱随机取3尾实验鱼, 即每组9尾。记录体长和体质量后, 使用肝素钠抗凝的注射器取血转入抗凝离心管中, 在4℃下以3500 r/min离心10min, 取上清血浆做好标记放置在-20℃冰箱保存, 用于血液学指标测定。取完血后进行解剖, 取肝脏、肠系膜脂肪、性腺、去除内脏后空壳称重计算肥满度(CF)、肝体比(HSI)、脏体比(VSI)、肠系膜脂肪系数(MFI)和性腺指数(GSI)。取0.3—0.5 g卵巢中后段保存于Bouin’s固定液中。固定后计数, 推算绝对繁殖力(F)和相对体重繁殖力(FW)。从每尾鱼固定好的卵粒中随机选择20粒利用显微镜镜台测微尺测定卵粒直径-卵径(ED)。同时取肝脏保存于-40℃冰箱,用于肝脏抗氧化能力的测定。
产后再培育养殖实验结束时, 停食24h, 记录每个网箱实验鱼剩余数量计算成活率。每个网箱随机取3尾实验鱼, 即每组9尾。测定性腺指数、绝对繁殖力、相对体重繁殖力及卵径与春季培育取样测定方法一致。将每组取完样各个组的剩余试验鱼放在相应暂养池暂养用来做人工繁殖, 暂养池水温27.9—28.2℃, 保持微流水状态。采用胸鳍基部两针法注射催产药物, 第1针: 促黄体生成素释放激素类似物(LRH-A2) 用量12 μg/kg; 第2针: LRH-A216 μg/kg, 脑垂体(PG) 4.5 mg/kg, 地欧酮8 mg/kg, 人绒毛膜促性腺激素(HCG)500 IU/kg。两针间隔12h。在12h效应时间前2—3h检查黄颡鱼雌性亲鱼状态, 待60%—70%的试验鱼个体能顺利排卵后, 即可以人工挤卵。采用“半干法”进行人工授精。所用精液是来自同1尾的瓦氏黄颡鱼, 进行人工授精。完成授精后将受精卵置于孵化桶进行流水孵化, 水流速度约0.2 m/s, 孵化桶水温26—28℃。待受精卵发育至原肠胚期计算受精率(FR) (约受精8h后), 每个组随机取样 3 次, 每次不少于200粒卵。在受精率测定后, 每组3个平行, 1个平行取200粒受精卵。受精卵放置于底部直径长22 cm、高35 cm的相同15个圆柱小桶内进行孵化。小桶上方有注水装置使桶一直保持流动水状态, 温度保持在28.5—28.8℃。待所有仔鱼出膜后统计每个桶内孵化率(HR)。
每个桶内随机选择20尾初孵仔鱼, 利用游标卡尺测定初孵仔鱼长度(BL)。测定结束保存在装有Bouin’s固定液的离心管之后利用显微镜镜台测微尺测定初孵仔鱼卵黄囊长(YL)。
待出膜的鱼苗发育至可平游时, 统计各个桶的畸形率(MR)。待鱼苗由内源性营养转为混合营养时, 能主动开口摄食统计桶内鱼苗存活率即出苗率(ER)。随后不投喂, 水温环境控制不变, 统计仔鱼3d成活率(TSR)。
1.4 指标测定及方法
指标计算公式:
成活率(SR, %)=存活尾数/初始尾数×100
肥满度(CF, g/cm3)=体质量(g)/体长3(cm)×100
肝体比(HSI)=肝脏重/体质量×100
脏体比(VSI)=鱼空壳重/体质量×100
肠系膜脂肪系数(MFI)=肠系膜脂肪重/体质量×100
性腺指数(GSI)=性腺重/体质量×100
绝对繁殖力(F, 粒)=(样品卵粒数目/样品重)×性腺重
相对体重繁殖力(FW, 粒/g)=绝对繁殖力/体质量
受精率(FR, %)=受精卵数/总卵数×100
孵化率(HR, %)=孵化出苗数量/受精卵数量×100
畸形率(MR, %)=畸形苗数量/孵化出苗数量×100
出苗率(ER, %)=出苗数量/受精卵数量×100
仔鱼3d成活率(TSR, %)=3d 仔鱼存活数量/孵化出苗数量×100
肝脏抗氧化酶活性及血浆生化指标采用南京建成生物工程研究所试剂盒测定。具体测定步骤按照说明书进行。利用细胞破碎仪制作肝脏组织匀浆, 组织匀浆中蛋白质浓度及血浆蛋白浓度采用南京建成BCA微板法。
1.5 数据处理
数据采用SPSS 26.0软件进行单因素方差分析(One-way ANOVA), 利用Duncan’s多重比较确定组间差异的显著性,P<0.05时表示在统计学上差异显著。文中数据均用平均值±标准差(mean±SD)表示。
2 结果
2.1 饲料中小麦胚芽对黄颡鱼雌性亲鱼肝脏抗氧化性能的影响
饲料中小麦胚芽添加对黄颡鱼雌性亲鱼肝脏抗氧化性能有明显影响, 肝脏抗氧化相关酶活性及丙二醛(MDA)含量对小麦胚芽添加有明显响应。超氧化物歧化酶(SOD)活性随小麦胚芽含量增加呈先升高后下降的趋势(图1A), 在W10组达到最高(30.52±1.97) U/mg prot。W5组(23.47±2.65) U/mg prot、W10组(30.52±1.97) U/mg prot、W15组(25.31±2.36) U/mg prot和W20组(24.01±2.83) U/mg prot均显著高于W0组(7.20±2.10) U/mg prot。
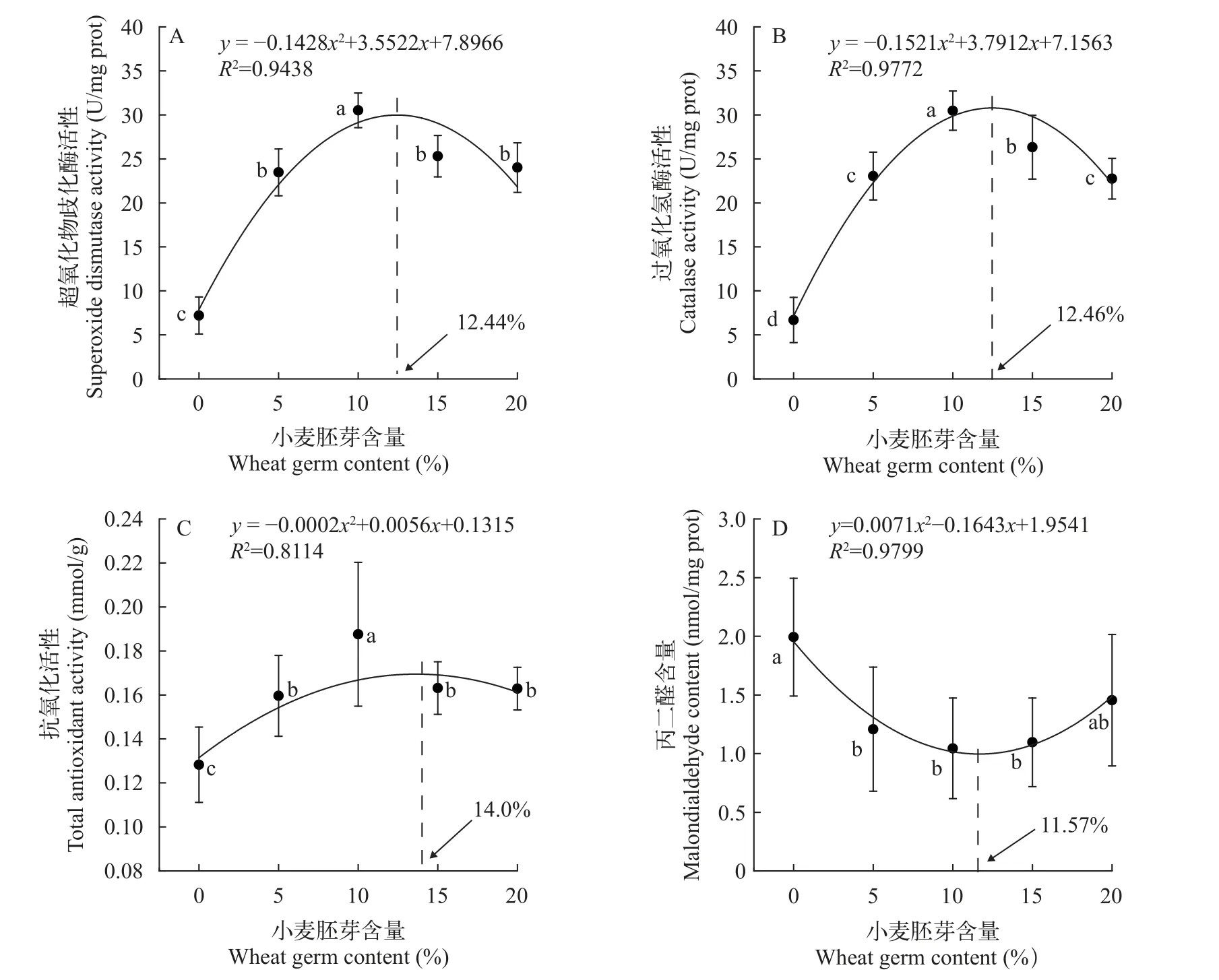
图1 小麦胚芽对黄颡鱼雌性亲鱼肝脏抗氧化能力的影响Fig.1 Effects of wheat germ on liver antioxidative capacity of female broodstock in yellow catfish
过氧化氢酶(CAT) 活性随小麦胚芽含量增加呈先升后降的趋势(图1B), 在W10组达到最高, 为(30.48±2.22) U/mg prot。W5组(23.05±2.71) U/mg prot、W15组(26.32±3.62) U/mg prot和W20组(22.75±2.32) U/mg prot均显著高于W0组(6.69±2.57) U/mg prot。W15组显著高于W5组和W20组。
总抗氧化能力(T-AOC) 活性同样随小麦胚芽含量增加呈先升后降趋势(图1C), 在W10组达到最高, 为(0.19±0.03) mmol/g。W5组(0.16±0.02) mmol/g、W15组(0.16±0.01) mmol/g和W20组(0.16±0.01) mmol/g均显著高于W0组(0.13±0.02) mmol/g, 但这3组之间无显著性差异(P>0.05)。
丙二醛含量则随小麦胚芽含量增加呈先降低后趋于稳定的趋势(图1D), W5组(1.21±0.53) nmol/mg prot、W10组(1.04±0.43) nmol/mg prot和W15组(1.10±0.38) nmol/mg prot显著低于W0组(1.99±0.50) nmol/mg prot。
根据小麦胚芽含量与抗氧化能力数据所拟合的多项式回归曲线显示, 不同抗氧化活性指标达到最大值时对应的小麦胚芽含量较接近。超氧化物歧化酶对应小麦胚芽含量为12.44%, 过氧化氢酶对应小麦胚芽含量为12.46%, 总抗氧化能力对应小麦胚芽含量为14.0%, 而与抗氧化性负相关的丙二醛含量在小麦胚芽含量为11.57%为最低(图1), 因此可得出小麦胚芽含量在11.57%—14.0%时肝脏抗氧化能力可达最大。
2.2 小麦胚芽对黄颡鱼雌性亲鱼血浆生理生化指标的影响
经过春季产前培育, 小麦胚芽对黄颡鱼雌性亲鱼血浆生理生化指标影响如表2所示。血浆中葡萄糖(Glu)含量随小麦胚芽含量增多出现波动, 在W5组达到最大值。在W10组下降为最小值, W10与W0组无显著性差异(P>0.05)。W15、W20组比W10组显著性增高。

表2 小麦胚芽对黄颡鱼雌性亲鱼血浆生理生化指标的影响Tab.2 Effects of wheat germ on plasma physiological and biochemical indexes of female broodstock in yellow catfish
高密度脂蛋白胆固醇(HDL-C)含量随小麦胚芽含量的增多呈先升后降趋势, W0组数值最小, 在W10组达最大值。随后开始下降, 但W15和W20组仍显著高于W0组。
低密度脂蛋白胆固醇(LDL-C)含量随小麦胚芽含量增多呈先升高后趋于稳定的趋势, 其余各组显著低于W0组。而4个组之间无显著差异(P>0.05)。低密度脂蛋白胆固醇含量与高密度脂蛋白胆固醇含量的比值(LDL-C/HDL-C)随小麦胚芽含量增多显著下降, 其中W10组为最低, 显著低于W0、W15和W20组。小麦胚芽添加对实验组碱性磷酸酶(AKP)、甘油三酯(TG)、总蛋白(TP)、白蛋白(ALB)和总胆固醇(T-CHO)含量均未造成显著性影响(P>0.05)。
2.3 小麦胚芽对黄颡鱼雌性亲鱼成活率的影响
饲料中小麦胚芽的添加, 能有效提高黄颡鱼雌性亲鱼成活率(图2)。春季产前培育结果如图2A所示, W0组成活率最低(76.97±1.05)%, 小麦胚芽添加组成活率均显著高于W0组(P<0.05), W5组(89.70±2.10)%、W10组(89.70±1.05)%、W15组(88.48±4.58)%和W20组(91.52±7.35)%的成活率分别较对照组W0组提高了16.5%、16.5%、15.0%和18.9%(P<0.05), 但W5、W10、W15和W20 四组之间无显著性差异(P>0.05)。根据小麦胚芽含量与产前培育成活率实验数据拟合的多项式回归曲线显示, 在小麦胚芽含量11.99%时成活率达到最大值。
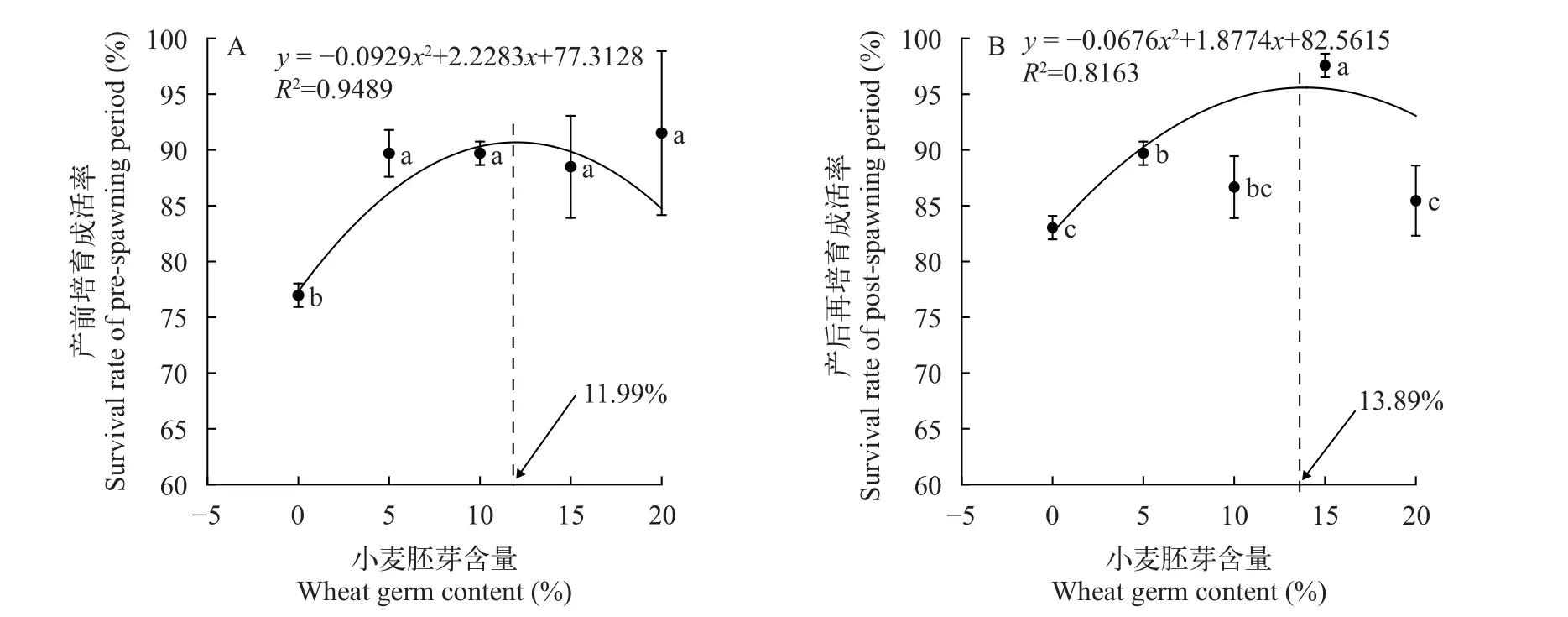
图2 小麦胚芽对黄颡鱼雌性亲鱼产前培育和产后培育成活率的影响Fig.2 Effects of wheat germ on survival rate of female broodstock in yellow catfish during pre-spawning and post-spawning period
产后再培育结果如图2B所示, W15组(97.58±1.05)% 成活率最高, 比W0组(83.03±1.05)%、W5组(89.70±1.05)%、W10组(86.67±2.78)%和W20组(85.45±3.15)% 显著提高了17.1%、8.8%、12.6%和14.2%。W5组显著高于W0、W20组, W0组、W10组和W20组之间无显著性差异(P>0.05)。根据小麦胚芽含量与产后再培育成活率实验数据拟合的多项式回归曲线显示, 在小麦胚芽含量13.89%时成活率达到最大值。
2.4 饲料中小麦胚芽对黄颡鱼雌性亲鱼形体指标和繁殖性能的影响
饲料中小麦胚芽对黄颡鱼雌性亲鱼形体指标的影响如表3所示, W5组的肠系膜脂肪系数显著高于W0组, 其余实验组之间无显著差异(P>0.05)。各实验组的肝体比、脏体比和肥满度指标之间均无显著性差异(P>0.05)。
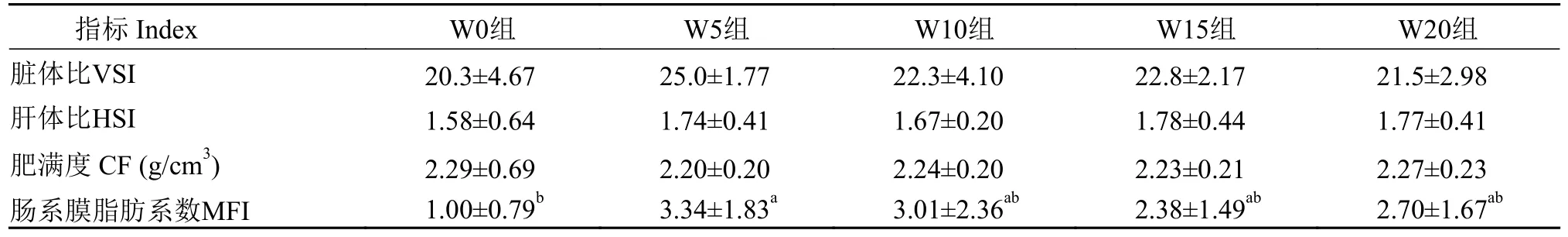
表3 小麦胚芽对黄颡鱼雌性亲鱼形体指标的影响Tab.3 Effects of wheat germ on physical indexes of female broodstock in yellow catfish
表4和表5分别展示了产前经过49d养殖和产后经过30d的营养强化培育后小麦胚芽对黄颡鱼雌性亲鱼繁殖性能的影响。小麦胚芽没有对试验组的性腺指数、绝对繁殖力、相对体重繁殖力和卵径产生显著性影响 (P>0.05), 但是绝对繁殖力和相对体重繁殖力与对照W0组相比有所提升。

表4 小麦胚芽对黄颡鱼雌性亲鱼产前培育繁殖性能的影响Tab.4 Effects of wheat germ on spawning ability of female broodstock in yellow catfish during pre-spawning

表5 小麦胚芽对黄颡鱼雌性亲鱼产后再培育繁殖性能的影响Tab.5 Effects of wheat germ on reproductive performance of female yellow catfish after post- spawning rebreeding
在产后再培育阶段(表6), 小麦胚芽添加量超过10%后, 黄颡鱼人工繁殖受精率显著下降(P<0.05),但W5、W10和W15组畸形率显著低于对照组(P<0.05),孵化率也略高于对照组(P>0.05)。综合受精率、孵化率和畸形率结果, 小麦胚芽在产后在培育阶段添加量不应超过10%。
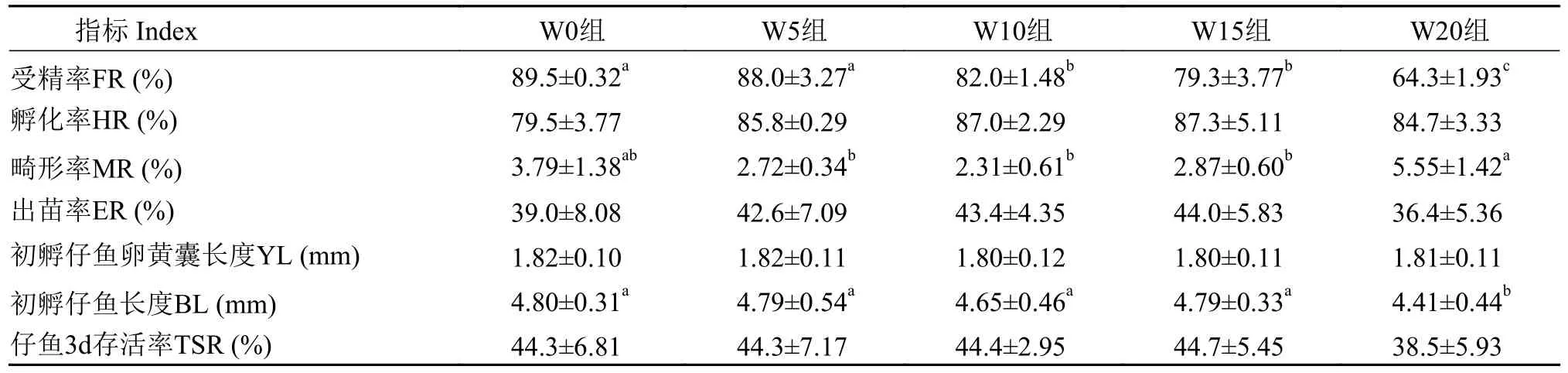
表6 小麦胚芽对产后再培育黄颡鱼雌性亲鱼繁殖性能及仔鱼质量的影响Tab.6 Effects of wheat germ on quality of eggs and larvae of female yellow catfish after post- spawning rebreeding
3 讨论
3.1 小麦胚芽对黄颡鱼雌性亲鱼肝脏抗氧化能力的影响
肝脏不仅是机体生理代谢的中心器官, 还能合成卵巢发育所需的卵黄蛋白原[20,21]。机体的抗氧化系统能保护机体免受活性氧增加导致的氧化损伤, 而肝脏的抗氧化系统对环境和营养变化较敏感。抗氧化系统包含有超氧化物歧化酶和过氧化氢酶等一系列抗氧化酶[22], 而丙二醛是脂质过氧化的产物, 含量与机体的健康呈负相关[23]。总抗氧化能力活力可以作为全面评价亲鱼抗氧化性能的依据[24], 能有效抵御自由基从而保障机体正常生理功能[25]。本次实验结果中肝脏抗氧化酶活性的显著提高, 脂质过氧化的风险被降低, 这与Mutlu等[26]在果蝇和El-Sisy等[12]在雄兔中饮食中添加小麦胚芽结果一致。现代鱼类养殖和亲本培育过程中, 主要以饲料为主, 过量投喂会导致肝脏和消化系统出现不同程度的损伤, 而小麦胚芽添加能改善肝脏抗氧化性能, 从而改善卵黄蛋白原合成状况, 这可能是其提高繁殖性能的重要原因。
研究表明小麦胚芽具有丰富的黄酮类化合物、维生素E、维生素C、类胡萝卜素和胚芽多糖,能清除多余的氧自由基, 降低脂质被氧化的风险及增强还原性, 通过与金属离子发生配位效应提高超氧化物歧化酶、过氧化氢酶抗氧化酶的活性来提高抗氧化能力[8,27,28]。关于小麦胚芽抗氧化的功效是否能在鱼类中体现出来, 较少有研究报道, 在亲鱼上的作用更是较少有关注。本实验通过肝脏抗氧化酶活性指标的测定, 确定了小麦胚芽可以作为抗氧化剂和自由基清除剂[29], 提高黄颡鱼雌性亲鱼肝脏抗氧化能力, 肯定了使用小麦胚芽后对黄颡鱼雌性亲鱼肝脏健康的积极作用, 为小麦胚芽在鱼类亲本的科学培育方面提供了重要依据。
3.2 小麦胚芽对黄颡鱼雌性亲鱼血浆生理生化指标的影响
鱼类自身新陈代谢及健康状态会反映在血浆生理生化指标中, 而指标与饲料营养关系紧密[30]。鱼类吸收利用饲料中碳水化合物能力较差, 黄颡鱼属杂食性鱼类, 杂食性和植食性利用碳水化合物能力高于肉食性鱼类[31]。小麦胚芽中含有较高的碳水化合物, 本实验结果中W10组血糖水平与W0组无显著性差异(P>0.05), W15和W20组虽显著升高但趋于稳定, 结合所在组实验鱼的成活率, 说明黄颡鱼雌性亲鱼能够很好地消化吸收并利于小麦胚芽中的碳水化合物作为能量来源之一, 且在10%含量时达到更好的利用度, 可以作为蛋白质和脂肪等主要能量来源的补充。
高密度脂蛋白胆固醇负责将组织胆固醇运输至肝脏进行清除, 含量与健康状况呈正相关[32]。低密度脂蛋白胆固醇负责将肝内胆固醇输出至组织中, 含量与健康状况呈负相关[32]。低/高密度脂蛋白胆固醇含量比值指标用来反映机体是否存在潜在血脂异常情况, 比值与健康呈负相关[33]。本实验结果显示, 随着小麦胚芽含量增多, 虽然低密度脂蛋白胆固醇和高密度脂蛋白胆固醇含量均上升, 但高密度脂蛋白胆固醇含量上升更为迅速, 因而低/高密度脂蛋白胆固醇含量比值下降, 利于健康, 尤其在W10组效果最优。这说明小麦胚芽对黄颡鱼雌性亲鱼的血脂代谢和血脂健康有利, 建议小麦胚芽添加含量为10%。本研究结果与Reza Zakeri等[34]、Rezp等[35]在人类研究中结果一致, 推测其原因也类似, 学者认为小麦胚芽中含有黄酮类化合物、植物甾醇、纤维及其蛋白成分能促进脂肪代谢, 防止脂肪过度沉积[35—37]。此外, 本研究中血浆中总蛋白、白蛋白和碱性磷酸酶没有显著变化, 说明小麦胚芽给黄颡鱼雌性亲鱼的肝脏未造成负面影响。
3.3 小麦胚芽对黄颡鱼雌性亲鱼成活率、形体指标及繁殖性能的影响
小麦胚芽对黄颡鱼雌性亲鱼的成活率有较大提升, 这与小麦胚芽使用在对虾[15]、鲈[16]上结果不同, 原因可能是其作用在对虾、鲈幼鱼阶段, 而本实验作用于亲鱼, 且实验设计基于池塘养殖大水体环境, 更具有实际生产参考价值。春季产前培育阶段, 小麦胚芽添加组均能显著提高雌性亲鱼的成活率, 推测与小麦胚芽中富有的维生素群体、不饱和脂肪酸和多种生物活性物质有关, 不仅利于雌性亲鱼的血脂代谢, 提高了机体的营养水平, 还通过抗氧化能力的提升, 以及不饱和脂肪酸的补充, 对亲鱼存活和繁殖起到积极作用。产后再培育阶段,W5组和W15组也显著提高了黄颡鱼雌性亲鱼的成活率, 且效果更明显, 推测由于产卵后的雌性亲鱼体能耗损严重, 比春季培育的雌性亲鱼更虚弱, 此时用小麦胚芽增强免疫力效果显著, 能够帮助其恢复健康。但是小麦胚芽添加量要适中, 碳水化合物水平过高可能会对繁殖产生负面影响。
脂肪成本低, 且容易被鱼类吸收利用, 是鱼类较为理想的能量来源。鱼类需要一定的脂肪蓄积预防外界食物缺乏等不利情况的出现, 雌性亲鱼性腺发育与成熟更是需要大量脂肪积累, 以满足卵子正常发育的需求。脂肪主要蓄积在肝脏、肠系膜和肌肉等部位, 适度蓄积脂肪的现象属于机体营养良好的表现[38]。但脂肪过度沉积, 会造成免疫力下降和脂毒性等不良影响[38]。黄颡鱼会最先在肠系膜上蓄积脂肪, 胡伟华等[39]研究发现, 适量的肠系膜脂肪才能保证黄颡鱼雌性亲鱼繁殖性能及鱼苗质量。本研究春季产前培育阶段, W0组的肠系膜脂肪堆积相比于其他实验组较少, 同时W0组的成活率也是最低, 说明肠系膜脂肪过少不利于其开春后的存活, 不能满足繁殖期黄颡鱼雌性亲鱼的生理需求。而小麦胚芽添加组中肠系膜脂肪系数数值最高的也在适宜的范围内(3.34%), 且亲本成活率高, 同时各实验组的肥满度、肝体比和脏体比指标均无显著差异, 表明适当的小麦胚芽添加量, 不仅能提高黄颡鱼雌性亲鱼营养水平, 且不会造成过度脂肪沉积, 使鱼肥满度适中。在欧洲鲈[16]、半滑舌鳎[17]上的研究表明, 小麦胚芽对幼鱼的肥满度、肝体比和脏体比指标无显著影响, 并且适当的添加还可以提高蛋白质效率和饲料利用率。综上, 小麦胚芽合理添加量对鱼类不同发育阶段均会产生积极影响。
Greenhill[40]在临床采用小麦胚芽治疗不孕或者习惯性流产的女性, 效果显著, 其认为是小麦胚芽中丰富的维生素E、维生素A发挥了作用。Blanco等[41]学者认为, 烟草芽虫能利用小麦胚芽所拥有的均衡蛋白从而保证繁殖率。后续学者发现食用小麦胚芽能提高雄兔的繁殖性能[12], 认为是小麦胚芽中的维生素E、硒提高睾丸激素水平, 增加精子数量, 同时黄酮类化合物、维生素C和类胡萝卜素等能减少脂质过氧化。在本次产前培育和产后强化营养培养后, 小麦胚芽添加对绝对怀卵量和相对体重怀卵量均未产生统计学上的显著性影响, 这与郑紫薇等[42]、Blanco等[41]在鸟类和昆虫中研究结果一致, 与El-Sisy等[12]和Goane等[14]在哺乳和昆虫类结果不同, 可能与实验中取样存在个体差异有关。黄颡鱼属分批产卵鱼类, 性腺发育存在较大的个体差异性, 因此后续研究将增大样品量, 从而减少因样品量少引起的组内差异。
产后再培育阶段, 小麦胚芽W10、W15和W20组较对照组受精率下降, 可能是小麦胚芽的油脂80%为不饱和系, 其中n-6多不饱和脂肪酸(亚油酸)约占60%[8], 而鱼卵对n-3多不饱和脂肪酸的需求量要高于n-6系[43], 造成n-3多不饱和脂肪酸积累不够。但适宜小麦胚芽添加能提高孵化率, 降低鱼苗畸形率, 有利于实际生产。尽管如此, 本研究中小麦胚芽添加组绝对怀卵量和相对体重怀卵量平均值均高于未添加组, 尤其W15组绝对怀卵量和相对体重怀卵量均高于W0组30%以上, 这对实际生产将产生巨大的积极作用。
4 结论
黄颡鱼雌性亲鱼春季产前培育和产后再培育结果表明, 饲料中添加小麦胚芽能显著提高黄颡鱼雌性亲鱼肝脏抗氧化性能及营养水平。虽然饲料中的小麦胚芽对繁殖力没有直接显著性提升, 但是能显著提高亲本成活率, 从而提高雌性亲本有效利用率, 且能降低产后培育阶段雌性亲本人工繁殖子代畸形率, 有利于其规模化繁殖。综合考虑本研究结果, 建议在产前培育阶段的饲料中添加10%—15%小麦胚芽, 能够提升黄颡鱼雌性亲鱼的成活率、肝脏抗氧化性能和碳水化合物利用能力, 利于糖脂代谢和规模化繁殖。产后再培育阶段饲料中添加5%—10%小麦胚芽, 能提高亲本利用率。
猜你喜欢
农村百事通(2022年9期)2022-05-30
世界科学技术-中医药现代化(2021年5期)2021-11-05
青年文学家(2021年4期)2021-03-18
农民致富之友(2020年19期)2020-07-18
家庭百事通·健康一点通(2019年11期)2019-09-26
当代水产(2018年8期)2018-11-02
现代园艺(2018年2期)2018-03-15
渔业致富指南(2017年8期)2017-02-02
渔业致富指南(2016年7期)2016-03-28
湖南中医药大学学报(2015年1期)2016-01-06
